導(dǎo)語(yǔ):
看過(guò)無(wú)數(shù)篇凋亡小體相關(guān)的文獻(xiàn),倏然看到炎癥小體,是不是有些不適應(yīng)?炎癥小體的研究目前只有短短十幾年,但在近幾年已經(jīng)成為了研究熱門(mén)趨勢(shì)之一。如果你對(duì)細(xì)胞炎癥、細(xì)胞程序性死亡領(lǐng)域感興趣,炎癥小體是你不得不了解的關(guān)鍵詞。

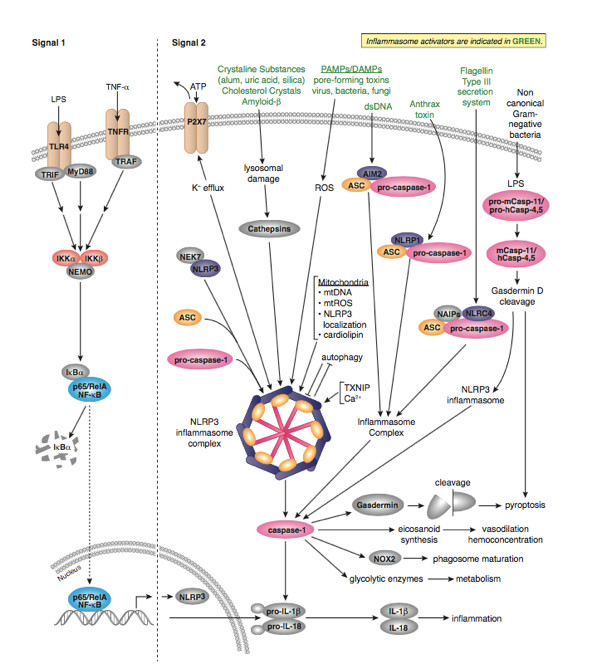
作為炎癥反應(yīng)的重要組成部分,炎癥小體在 2002 年被發(fā)現(xiàn)以后就一直是免疫和炎癥疾病領(lǐng)域的重點(diǎn)研究對(duì)象。炎癥小體(Inflammasome)是由胞漿內(nèi)模式識(shí)別受體 (PRRs) 參與組裝的多蛋白復(fù)合物,主要包含受體蛋白(即 PRRs)、接頭蛋白 ASC(也稱為 TMS1)以及下游的 Caspase-1。受體蛋白被激動(dòng)劑激活后,會(huì)隨之吸引 ASC 和 caspase-1 組裝成直徑可達(dá)微米級(jí)的炎癥小體,從而誘導(dǎo) caspase-1 自切割并活化。活性 caspase-1 一方面促進(jìn)包括白細(xì)胞介素-1β(IL-1β)和-18(IL-18)在內(nèi)的多種促炎性細(xì)胞因子的成熟和分泌,另一方面引發(fā)一種程序性炎性細(xì)胞死亡,即細(xì)胞焦亡(Pyroptosis)(圖 1)。
炎癥小體的受體蛋白包括 NOD 樣受體 (NLR) 家族成員(如 NLRP1, NLRP3,NLRC4 等)以及來(lái)自 HIN200 家族的 AIM2。NLR 家族成員是構(gòu)成炎癥小體的一大類受體蛋白,它們大多數(shù)含有 LRR(Leucine-rich Repeat)結(jié)構(gòu)域,可以識(shí)別細(xì)胞內(nèi)一系列的「危險(xiǎn)」信號(hào)。
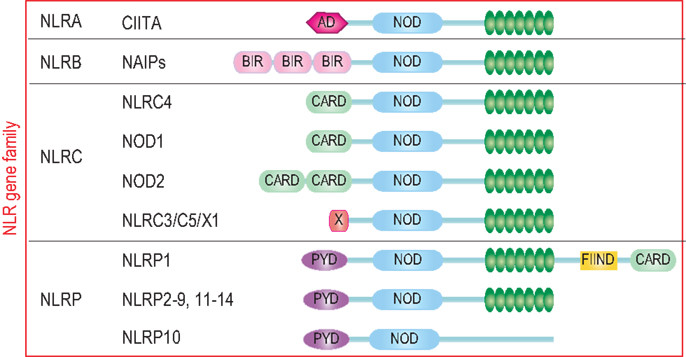
圖 2:NLR 家族成員結(jié)構(gòu)示意圖【1】
大部分炎癥小體只會(huì)被一種或少數(shù)幾種高度特異的激動(dòng)劑激活,但 NLRP3 是一個(gè)例外,它能夠?qū)?shù)量繁多并且在來(lái)源、化學(xué)組成、結(jié)構(gòu)性質(zhì)上都毫無(wú)關(guān)聯(lián)的激動(dòng)劑做出反應(yīng),因此它的功能范圍也是所有炎癥小體中最為廣泛的,是大家研究最多也最深入的一個(gè)代表。NLRP3 激動(dòng)劑不僅包括來(lái)自入侵病原體的病原相關(guān)分子模式(pathogen-associated molecular patterns,PAMPs)(例如細(xì)菌 RNA、細(xì)菌多肽類抗生素和流感病毒離子通道蛋白),還有受損機(jī)體自身產(chǎn)生的損傷相關(guān)分子模式(damage-associated molecular patterns,DAMPs)(例如 ATP),以及多種跟疾病密切關(guān)聯(lián)的分子(例如阿爾茨海默病中的β-淀粉樣蛋白(amyloid β)以及動(dòng)脈粥樣硬化中的膽固醇結(jié)晶(cholesterol crystal))等等。因此,NLRP3 炎癥小體在病原體感染、自身炎癥反應(yīng)、神經(jīng)退行性疾病、癌癥、2 型糖尿病等多種人類疾病中都發(fā)揮著舉足輕重的作用【2-7】。
NLRC4 炎癥小體特殊之處在于不含 PYD 結(jié)構(gòu)域而含有 CARD 結(jié)構(gòu)域,也就意味著其不需要 ASC 蛋白作為「橋梁」而可以直接通過(guò) CARD:CARD 相互作用和效應(yīng)蛋白連接。對(duì)細(xì)菌鞭毛蛋白和細(xì)菌保守的 type II 型分泌系統(tǒng) (TTSS) 的組分作出的應(yīng)答,NLRC4 (IPAF) 炎性體形成【8-12】。配體檢測(cè)和配體依賴型 NLRC4 寡聚化和炎性體激活需要【13,14】蛋白 NAIP 家族。在小鼠中,NAIP5 和 NAIP6 與鞭毛蛋白有關(guān),而 NAIP2 與 TTSS 桿蛋白發(fā)生相互作用【13,14】。NAIP 可識(shí)別人體內(nèi)的 TTSS 針蛋白 Cprl【14】。另外,PKCδ 對(duì)細(xì)菌感染作出應(yīng)答使 NLRC4 磷酸化,而合一磷酸化對(duì)炎性體組裝和 caspase-1 激活 是必要的【15】。
而 AIM2 則是一類 DNA 感受器,可以通過(guò)它 C 端 HIN200 結(jié)構(gòu)域識(shí)別并結(jié)合自體或異體的 DNA,這一過(guò)程是不依賴序列就能完成的。而 IFI16 炎癥小體被認(rèn)為可以在細(xì)胞核內(nèi)識(shí)別一些病毒如 KSHV 的 DNA,其結(jié)構(gòu)和 AIM2 類似,都需要 ASC 作為「橋梁」發(fā)揮連接作用【16-18】。
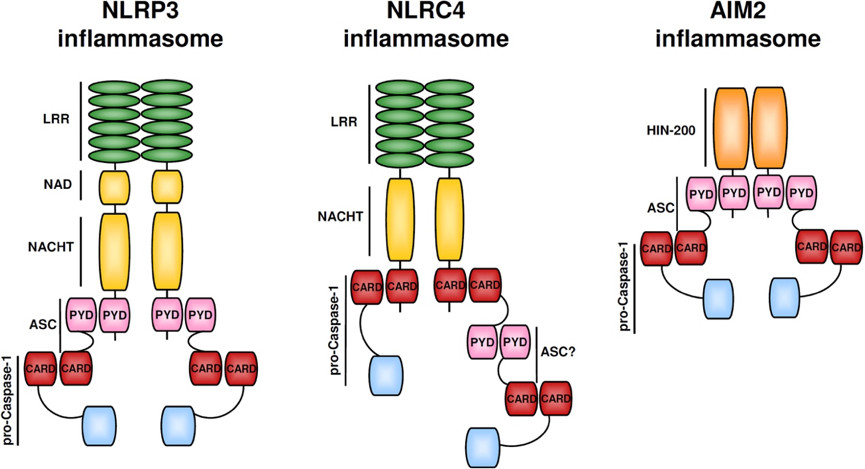
圖 3 幾種常見(jiàn)炎癥小體的結(jié)構(gòu)組成示意圖
另外,炎癥小體的組裝離不開(kāi)早期炎癥信號(hào)誘導(dǎo)的受體蛋白(如 NLR 家族成員,AIM2 等)、接頭蛋白 ASC/TMS1、pro-IL-1β 和 pro-IL-18 等分子的表達(dá)增強(qiáng),因此,大家常常會(huì)通過(guò) WB 來(lái)檢測(cè)這些分子的表達(dá)變化。
Caspase-1 又被稱為白介素-1? 轉(zhuǎn)化酶 (ICE/ICEα) ,是一種 I 類半胱氨酸蛋白酶。與 Caspase-1 功能接近的 Caspase 還包括 caspase-4、-5、-11 和 -12,它們一起構(gòu)成 Caspase-1 亞家族。Caspase-1 能夠?qū)⒀装Y細(xì)胞因子(如 pro-IL-1? 和干擾素-γ 誘導(dǎo)因子 (IL-18))裂解成成熟蛋白【19,20】。與其他半胱天冬酶一樣,caspase-1 通過(guò)酶原蛋白水解被激活,產(chǎn)生一個(gè)由兩個(gè)活性亞基(p20 和 p10)組成的四聚體。Caspase-1 具有巨大的氨基末端前體域,此區(qū)域包含一個(gè)半胱天冬酶募集區(qū)域 (CARD)。Caspase-1 過(guò)表達(dá)會(huì)誘導(dǎo)細(xì)胞凋亡【21】。Caspase-1 缺失小鼠在凋亡方面沒(méi)有嚴(yán)重缺陷,但在 pro-IL-1β 蛋白的成熟過(guò)程中有缺陷,而且這類小鼠可抵抗內(nèi)毒素休克【22,23】。研究發(fā)現(xiàn),caspase-1 至少有六種同工型,其中包括 caspase-1 α、β、γ、δ、ε 和 ζ (6)。多數(shù) caspase-1 同工型(α、β、γ 和 δ)生成的產(chǎn)物在 30-48 kDa 之間,并通過(guò)過(guò)表達(dá)誘導(dǎo)細(xì)胞凋亡。Caspase-1 ε 通常只包含 p10 亞基,不會(huì)誘導(dǎo)細(xì)胞凋亡,可作為一種顯性失活蛋白。Caspase-1 中廣泛表達(dá)的 ζ 同工型會(huì)誘導(dǎo)細(xì)胞凋亡,并且與 α 同工型相比,缺乏 39 個(gè)氨基末端殘基【24】。caspase-1 通過(guò)炎癥小體的寡聚化分子平臺(tái)激活,此中包含 caspase-5、Pycard/Asc 和 NALP1【25】。
Interleukin-1β (IL-1β) 是 caspase-1 的重要靶標(biāo)之一,也是一個(gè)參與許多免疫和促炎性反應(yīng)的多功能細(xì)胞因子【26】。它主要由激活的單核細(xì)胞和巨噬細(xì)胞產(chǎn)生。它通過(guò)各種接頭蛋白和激酶發(fā)出信號(hào),從而激活大量下游靶標(biāo)【27-31】。人 IL-1β 可以合成為 31 kDa 前體。為了獲得活性,前體必須在 Asp116 和 Ala117 位點(diǎn)之間被 caspase-1 剪切產(chǎn)生 17 kDa 成熟形式【32,33】。IL-1β 17 kDa 成熟形式的檢測(cè)是衡量 caspase-1 活性的一個(gè)很好指標(biāo)。

分析完以上炎癥小體研究相關(guān)靶點(diǎn),你是否想到要更加經(jīng)濟(jì)地購(gòu)買(mǎi)到該研究相關(guān)的多個(gè)靶點(diǎn)的抗體?CST 新推出了 Human Reactive Inflammasome Antibody Sampler Kit II #25620,可幫您探秘炎癥小體相關(guān)的科研問(wèn)題。這個(gè)產(chǎn)品包裝里包括:
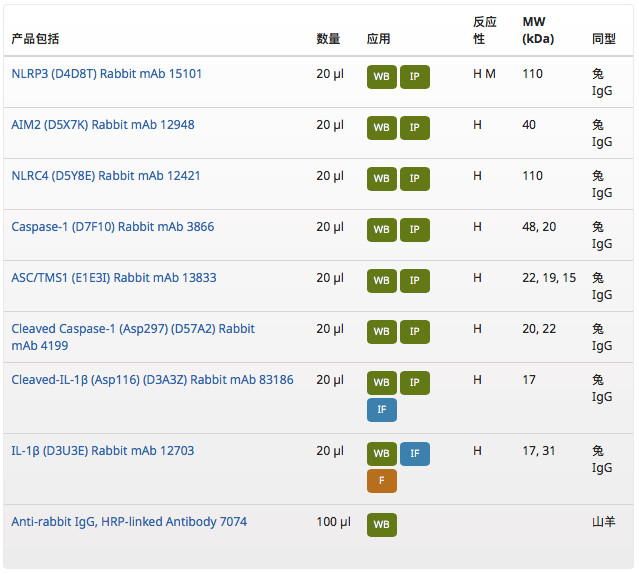
該產(chǎn)品的特點(diǎn):
- 用于檢測(cè)人樣品當(dāng)中的 NLRP3、NLRC4、AIM2、ASC/TMS1、Caspase-1(包含前體和切割活化型)、IL-1β(包含前體和成熟型)等多個(gè)分子;
- 小包裝抗體,經(jīng)濟(jì)實(shí)用;
- 非常適合于初期炎癥小體相關(guān)和 caspase-1 激活引起的炎癥的信號(hào)通路的探索。
小編點(diǎn)評(píng):
炎癥小體是細(xì)胞焦亡以及其激活 caspase-1 引起的 IL-1β分泌的炎癥反應(yīng)的重要調(diào)控因素。除此之外,Gasdermin D 也是細(xì)胞焦亡下游信號(hào)通路的重要靶點(diǎn),在各個(gè)領(lǐng)域中發(fā)揮的作用正亟待揭曉,也是目前研究的熱點(diǎn)之一。
提示:
CST 出品了一系列不同研究領(lǐng)域的小包裝抗體組合套裝,廣受大家歡迎,歡迎大家咨詢,了解更多。
參考文獻(xiàn):
- Zhong Y et al. Front Immunol. (2013) 4:333.
- Broz, P. and Dixit, V.M. (2016) Nat Rev Immunol 16, 407-20.
- Guo, H. et al. (2015) Nat Med 21, 677-87.
- Jo, E.K. et al. (2016) Cell Mol Immunol 13, 148-59.
- Rathinam, V.A. and Fitzgerald, K.A. (2016) Cell 165, 792-800.
- Shao, B.Z. et al. (2015) Front Pharmacol 6, 262.
- Schroder, K. and Tschopp, J. (2010) Cell 140, 821-32.
- Mariathasan, S. et al. (2004) Nature 430, 213-8.
- Poyet, J.L. et al. (2001) J Biol Chem 276, 28309-13.
- Franchi, L. et al. (2006) Nat Immunol 7, 576-82.
- Miao, E.A. et al. (2006) Nat Immunol 7, 569-75.
- Miao, E.A. et al. (2010) Proc Natl Acad Sci U S A 107, 3076-80.
- Kofoed, E.M. and Vance, R.E. (2011) Nature 477, 592-5.
- Zhao, Y. et al. (2011) Nature 477, 596-600.
- Qu, Y. et al. (2012) Nature 490, 539-42.
- Roberts, T.L. et al. (2009) Science 323, 1057-60.
- Hornung, V. et al. (2009) Nature 458, 514-8.
- Fernandes-Alnemri, T. et al. (2009) Nature 458, 509-13.
- Thornberry, N.A. et al. (1992) Nature 356, 768-74.
- Martinon, F. and Tschopp, J. (2004) Cell 117, 561-74.