

Hieff NGS? In-Situ DNA Binding Profiling Library Prep Kit for Illumina? V2
產品名稱: Hieff NGS? In-Situ DNA Binding Profiling Library Prep Kit for Illumina? V2
英文名稱: Hieff NGS? In-Situ DNA Binding Profiling Library Prep Kit for Illumina? V2
產品編號: 12597ES
產品價格: 詢價
產品產地: Yeasen
品牌商標: Yeasen
更新時間: 2025-11-07T19:45:03
使用范圍: null
- 聯系人 : 李自轉
- 地址 : 上海市浦東新區天雄路166弄一號樓三層南單元
- 郵編 : 200030
- 所在區域 : 上海
- 電話 : 139****5640 點擊查看
- 傳真 : 點擊查看
- 郵箱 : lizizhuan@yeasen.com
- 二維碼 : 點擊查看
Hieff NGS® In-Situ DNA Binding Profiling Library Prep Kit for Illumina® V2 是針對Illumina®高通量測序平臺研發的用于CUT&Tag實驗的文庫構建試劑盒,適用于100-100,000個細胞起始量的樣本建庫。CUT&Tag是研究蛋白與DNA互作的技術,相較于傳統的ChIP-seq、CUT&RUN,該技術具有文庫構建時長更短(僅需7小時)、操作更簡單、對起始樣本要求更低、抗體投入量更少、文庫產量更高等優點。經過細胞捕獲、一抗孵育、二抗孵育、轉座酶孵育、轉座酶激活、摻入DNA標準品、細胞裂解、磁珠回收gDNA、文庫擴增和磁珠分選等步驟,靶蛋白結合的DNA片段最終轉化為適用于Illumina®平臺測序的文庫。
本試劑盒包含兩個獨立模塊:BOX-I和BOX-II。BOX-I包含結合細胞的ConA Beads、提取DNA 的DNA Extract Beads以及文庫純化的DNA Selection Beads,BOX-II包含細胞透化劑、抗體結合buffer、轉座酶結合buffer、蛋白酶K以及后續文庫擴增所需的所有試劑。此外,本試劑盒已在不同種類細胞(如293T、K562、CHO、ESC細胞等)中進行了驗證,均具有良好的建庫效率和建庫產量。本試劑盒提供的所有試劑都經過嚴格的質量控制和功能驗證,最大程度上保證了文庫的穩定性和重復性。
- 純化磁珠升級:表觀實驗專用磁珠,更多小片段有效富集,結果更可靠。
- pA/G-Tnp升級:精準靶向,切割效率高,背景低。
- 實驗流程優化:簡化試劑配置,兼容更多樣本類型。
- 添加DNA Spike-in:數據標準化,減少實驗誤差,矯正組間差異。
- 應用方向:蛋白質-DNA互作研究:組蛋白修飾調控、轉錄因子調控等表觀作用機制研究
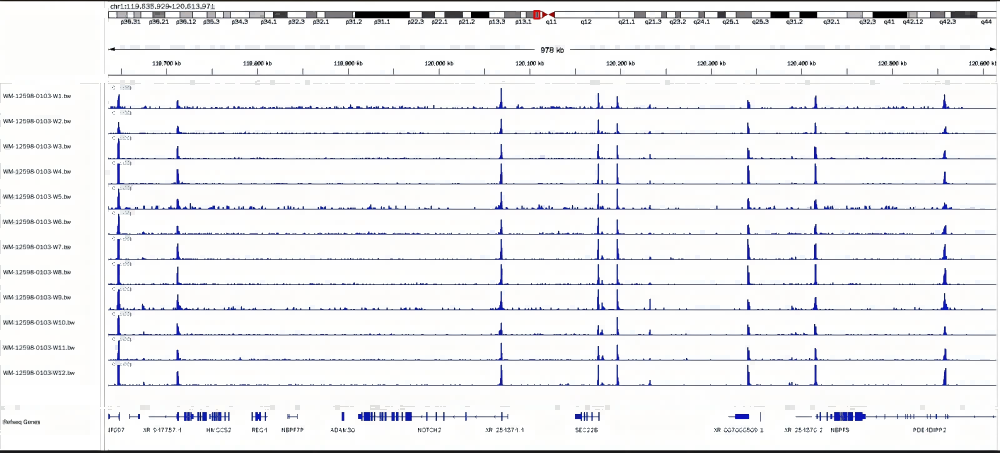
1. 不同投入量細胞實驗結果一致性較高
293細胞樣本投入細胞100個到10萬細胞數,使用H3K4me3抗體,100個到10萬個細胞樣本結果一致性較高。
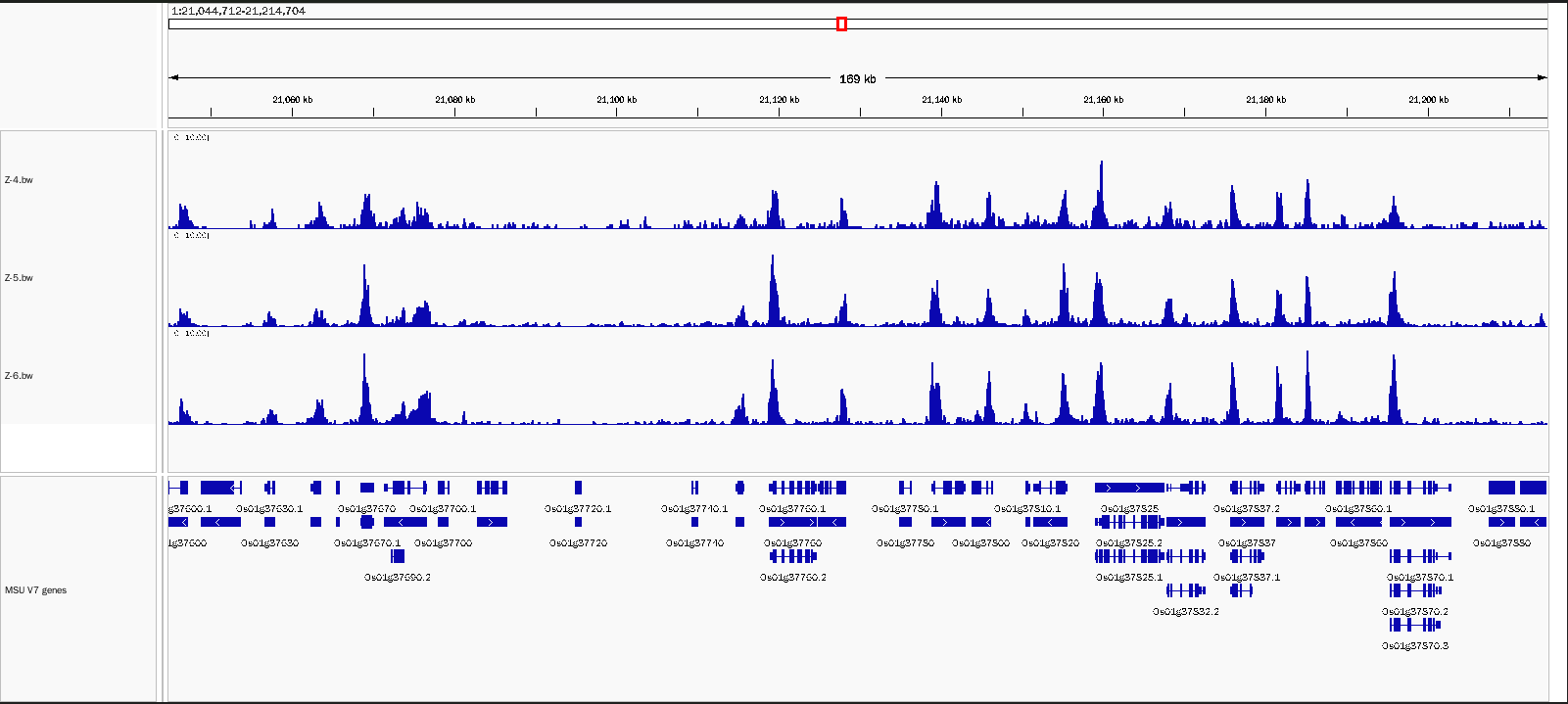
2. 適用植物細胞核樣本
選用水稻樣本進行細胞核抽提,5萬細胞投入量,使用H3K4me3抗體,樣本重復性好,背景噪音低。
BOX-I:2-8℃保存,BOX-II:-25~-15℃保存,有效期1年。
PartⅠ:產品定位相關
Q1.本產品是否適用于細菌等原核生物的蛋白核酸互作研究?
A1:原核生物沒有組蛋白,且有細胞壁,要做的話建議用 0.3%的甲醛交聯后破除細胞壁再去做,理 論上可行。
Q2.請問這個實驗能代替 ChIP 實驗嗎?
A2:對,完美代替 ChIP
Q3.這個可以用來篩選免疫受體所識別的病毒 DNA 區段嗎?
A3 :如果定位在細胞核里面應該可以做,在細胞質里面可能會由于 Digitonin 透化細胞膜導致 DNA 片段丟失。建議客戶可能先使用甲醛交聯,防止 DNA 丟失,再嘗試用本試劑盒建庫。
Q4.植物轉錄因子做 cut &tag 有什么限制?我們有個轉錄因子做ChIP 重復性很差,不知道做 CUT &tag 可不可行
A4 :ChIP 實驗不管是組蛋白修飾還是轉錄因子都存在重復性差、背景信號高的問題,CUT&tag 實驗 最大的優勢就在于實驗重復性好,實驗背景信號低。目前已經報道了非常多的樣本使用 CUT&Tag 技 術成功的案例,植物樣本實驗室制備原生質體或者細胞核實驗都行。
PartⅡ:產品實驗相關
Q5.在植物上融合什么標簽比較合適呢?
A5 :這個需要看客戶有什么標簽抗體,成功率主要取決于抗體是否好用以及蛋白表達豐度。盡量選 擇已經測試做 CUT&tag 或者 CUT&RUN 的抗體或者已經有相關文章報道的抗體進行實驗
Q6.回收 DNA 的磁珠有單賣的嗎
A6:本試劑盒中Box Ⅰ帶有 DNA 回收磁珠,客戶如需額外購買,本司可以提供(Cat#12601)
Q7.已知的轉錄因子,做 CUT&tag 實驗,是否可以不測序,直接 qPCR 檢測目的基因呢?
A7 :可以,但 qPCR 必須在文庫擴增之后做,我們的 spike-in 可以作為內參進行相對定量。
Q8.植物中怎么捕獲細胞?
A8 :植物細胞在使用本試劑盒建庫之前需要破除細胞壁或者直接提取細胞核,植物細胞核提取方法 可咨詢翌圣生物,我們有完整的植物細胞核提取方案。
Q9.對抗體的要求如何
A9 :抗體一般按質量標準分為免疫熒光級別、western blot 級別、免疫共沉淀(IP)級別和染色質免 疫共沉淀(ChIP)級別,CUT&tag 或 CUT&RUN 級別,本試劑盒要求的抗體盡量使用更高級別的抗 體。
Q10.回收 DNA 的磁珠可不可以用于做 ChIP 里面的回收 DNA 步驟?
A10 :可用回收,但是 ChIP 實驗一般捕獲下來的 DNA 太少,而且體積較大,利用磁珠回收可能會有 損失。建議酚氯仿抽提后沉淀,損失會小一些。
Q11.這個實驗怎么做重復?平行重復還是獨立重復?
A11 :NGS 數據一般是兩到三次平行重復。
Q12.想請問一下之前已經甲醛交聯過的細胞 還可以做 CUT&tag 嗎?
A12 :甲醛交聯的細胞可以繼續做,但甲醛濃度過高交聯時,后續需要增加解交聯步驟。
Q13.conA 磁珠用完了,其他組分還有,conA 磁珠有單賣嗎?或者有替代品嗎?
A13 :19810ES
Q14.spike in mix 具體的作用是怎樣的呢 ?
A14 :spike in 主要用于對給實驗數據定量提供依據,類似于內參的作用,可以看我們的PPT 上有展 示。對于不知道靶位點的目標蛋白或者處理條件會造成細胞內目標蛋白與 DNA 結合能力大幅度改變 的情況,spike in 能夠幫助做出更準確的定量,反映這些情況造成的實驗假陰性現象。此外,我們的 spike in 可以用于對目標 DNA 片段進行絕對定量,保證文章數據的準確性。
Q15.是不是需要先構建標簽載體,轉入原生質體再做后續的?還是用轉基因植株做?
A15 :原生質體可以直接用本試劑盒進行實驗,植株要先提取細胞核,詳情同問題 5 。若有植物靶蛋 白對應的抗體,可直接實驗,若無對應抗體,需要構建轉基因植株后使用標簽抗體實驗。
可以。
Q16. 請問有做過楊樹的嗎?
A16: 目前內部暫未有專門針對楊樹物種測試,但理論上是可行的。
Q17.測序數據中的背景噪音,主要指的哪一塊,不同的樣本有相對的參考么,還是越低越好?
A17 :背景噪音主要來源于抗體的非特異性結合,可以根據文庫分布情況以及生信分析的指標判斷背 景噪音。
Q18.轉基因材料帶有 flag 標簽就可以直接做嗎?
A18 :可用直接做。
Q19.做 cut&tag 總是 duplicate 多該怎么優化呢
A19 :可以適當提高細胞投入量和降低 PCR 循環數
Q20.之前做western flag 標簽有非特異條帶,這樣還能做嗎?
A20 :非特異性條帶做 CUT&Tag 可能會產生非特異性的結合位點,建議客戶更換抗體。如果非特異 性條帶不是 DNA 結合蛋白也可以試一下
Q21.IgG 對照也不用做嗎?
A21 :建議做的時候帶一個 IgG 對照來評判結合特異性。
Q22. 細胞捕獲后直接破膜加抗體,后續的孵育切割需要幾個小時的時間,這時細胞內的轉錄因子的 結合會不會產生變化?理論上說,轉錄因子對外界信號刺激很敏感.
A22:我們使用的緩沖液都是貼近細胞真實狀態的緩沖液,影響會比較小。當然,如果擔心自己的靶 蛋白和一個結合較弱的轉錄因子在建庫過程中與 DNA 解離了,可以用0.2%的甲醛輕微交聯一下。
Q23. IgG 和不加一抗都是背景很高
A23:很有可能是實驗過程中,pAG-Tn5 與抗體偶聯步驟出現問題,導致 tn5 酶非特異性切割。
Q24.請問打孔之后細胞是否已經失活,與 DNA 結合的蛋白質是否不在脫落?
A24:實驗過程中會添加 digitonin 進行細胞打孔,主要作用是為了使抗體以及 pAG-tn5 等大分子進 入細胞核內進行抗體靶向識別及酶切,不會使細胞失活。
Q25. CUT&tag 做組蛋白修飾用的都是通用抗體嗎 ?會不會對物種有選擇
A25:抗體并非都是通用抗體,同一支抗體并非能識別所有物種,所以實驗時,需要根據自己的樣本 類型進行測試實驗,選取更適合自己樣本的抗體。
Q26. 二抗的選擇有特殊的要求嗎?
A26:首先,二抗需要選擇與一抗來源對應,如一抗為兔源抗體,二抗需選擇抗兔的二抗,另外,建 議選擇含抗體重鏈和輕鏈的二抗,并且盡量不要偶聯其他標記物如 HRP 、FITC 等。
Q27. 貼壁細胞實驗準備時,能使用胰酶消化么?
A27:我們比較建議使用胰酶消化,雖然有地方報道使用胰酶會破壞細胞表明糖蛋白從而破壞細胞與 ConA 的結合,但是使用細胞刮進行刮取時,很容易壓迫甚至裂解細胞,造成后續的一些實驗背景干 擾。
PartⅢ:產品技術相關
Q28.CUT&Tag 一般適用于哪些類型蛋白質的研究?能否應用于異染色質上的位點?
A28:與 ChIP-seq 相同,CUT&Tag 主要用于分析全基因組范圍內的 DNA-蛋白質結合位點分析和組蛋 白修飾研究,包括存在于異染色質區域的蛋白,如存在于染色質壓縮纏繞區的 H3K27me3。當用于研 究轉錄因子時, 轉錄因子的豐度較低會導致其文庫產量也相比組蛋白修飾研究的文庫產量低,但同 樣可以獲得好的測序及數據分析結果。
Q29.CUT&Tag 適用的細胞類型有哪些?
A29:1)常見哺乳動物細胞 CUT&Tag 在常見哺乳動物細胞系上的應用比較成熟,常見的懸浮細胞系 (如 K562)正常離心收集棄去 培養基即可使用。對于大部分貼壁細胞系來說, 可按照常規流程使用 胰蛋白酶消化后得到細胞懸液后可正 常進行 CUT&Tag 實驗。需要注意的是,消化后的細胞由于表面 糖蛋白減少,與 ConA 磁珠的結合能力有所 下降,可能會損失 10-20%的細胞。建議可采用刮下的辦 法得到懸浮細胞。 另外理論上,CUT&Tag 可以在貼壁細胞中直接使用。ConA 磁珠的主要作用是方 便換液并保持細胞呈分 散狀態,鑒于 CUT&Tag 基本邏輯類似于胞內流式或免疫熒光,因此可以參考 免疫熒光的操作路線,用貼壁 較牢固的細胞或適度固定的細胞嘗試 CUT&Tag 實驗。
2)植物及其他細胞 可采用溫和的方式制備原生質體或者提取細胞核進行實驗。需要注意的是, 細胞 核與 ConA 磁珠的結合率為~50-80%,完整細胞與 ConA 磁珠的結合率>90% 。 另外理論上,植物細胞 按照免疫熒光進行染色方案可以直接調整到 CUT&Tag 實驗,其他物種邏輯上類似。
Q30.在準備實驗細胞樣品時,對細胞的狀態有什么具體要求?
A30:準備實驗細胞樣品時,對細胞的狀態有什么具體要求? 在準備細胞時需要保證細胞處于正常的 生長狀態,生長狀態不佳的細胞,其胞內的蛋白核酸結合相互 作用狀況會發生改變,而死亡細胞的目 的蛋白可能從染色質上的脫離,甚至細胞和染色質的破碎降解,嚴 重影響實驗結果。可以在準備細胞 樣品時用臺盼藍染色來鑒定細胞活性,細胞形態正常且樣品活細胞占 >90%即可。 對于凍存的樣本, 相對于 LN2 快速凍存的樣本,更加傾向使用“Mr. Frosty”緩慢凍存的樣本。冰凍切片理論上可以直接 在切片上進行 CUT&Tag 實驗;事先甲醛固定的石蠟切片也有可能直接進行 CUT&Tag 研究。操作時 按照免疫熒光的操作方式進行。
Q31.如何選擇用于 CUT&Tag 的抗體?
A31 :一抗建議使用 ChIP 級別的抗體;對于二抗,則選擇與 proteinA/G 親和力高的抗體(絕大部分物 種都可以)。
Q32.在實驗過程中,觀測到磁珠有凝聚現象怎么辦?
A32:在孵育時間比較長,特別是 4℃過夜的情況下,可能會出現部分磁珠在溶液內凝聚。一般情況下 只要大 部分磁珠仍處于溶液浸潤范圍內,干結在溶液邊緣和液面上方的磁珠比例不大,則仍然能夠 得到正常的實驗 結果,繼續進行后續實驗即可。 盡管如此,仍需注意高轉速離心或過長時間置于磁 力架,以及磁珠脫離液相時間過長而造成磁珠干結可 能會加劇磁珠的凝聚。在實驗過程中應盡量避 免此類操作,以免為實驗結果帶來不確定的影響。
Q33.對于實驗流程中可選的溫度條件和時間,如一抗孵育和細胞裂解消化的步驟,該如何進行選擇?
A33 :在一抗孵育步驟中,對于組蛋白修飾靶點,或者豐度較大的 DNA 結合蛋白靶點,如 RNA PoI II,在室溫下 孵育兩小時已經完全足夠。而對于豐度較低的轉錄因子則推薦 4ºC 孵育過夜。 在細胞 裂解后加入 ProteinaseK 消化的步驟中,55℃ 1h 小時和 37℃過夜并無特別大的區別。通常情況下 55℃ 消化 1h 小時即可,較高的反應溫度也有利于破壞胞內 DNase 的活性。
Q34.對于 DNA 擴增和建庫相關的試劑,如引物和擴增酶,能否使用其他來源的同類試劑?
A34:不建議使用本試劑以外的試劑混搭進行實驗。整個說明書的實驗流程,包括 PCR 程序都是針對 試劑盒內部所 提供的試劑進行優化的,引入任何非本試劑盒的試劑都可能會造成實驗的失敗。
Q35.對于 CUT&Tag 的實驗樣本,二代測序上機應該如何選擇?有何特殊的要求?
A35 :推薦常規的雙端 150-300bp 測序,如常見 HiSeq 系列。
Q36.對于已經完成的 CUT&Tag 的實驗,如何檢驗實驗結果?
A36 :二代測序標準的樣檢方法:利用 Qubit 熒光計、安捷倫 2100 可以檢定文庫的產出和片段分布。 合格的文庫應該可見類似 ATAC-seq 的典型梯狀條帶分布,而文庫的產量則會因為具體靶蛋白的豐度 不同而不同。若已知靶點結合序列的相關信息,也可設計相應位點的 PCR 引物與 GAPDH 一類的內 參位點對照,通過 qPCR 來確定目的靶點的富集程度。 粗略的實驗結果評估可以通過適當增加 PCR 循環數后利用瓊脂糖電泳來確定實驗是否成功。若同時進行了普通 IgG 替換目的抗體作為一抗的陰 性對照組,則可以在電泳圖上看到陽性組和陰性組不同的條帶圖形。過度的 PCR 會造成大 量的 PCR depulication 而帶來大量無效數據,因此也不可以用此類樣本進行回收建庫上機。 測序數據可以用 deeptools 的 plotHeatmap 或 plotProfile 來判斷富集效果。
Q37.CUT&Tag 目標片段富集的不夠好是什么原因?如何改進?
A37 參考免疫熒光的經驗,CUT&Tag 出現特異性不好的主要原因包括:二抗、轉座體(加鎂離子之 前)洗滌不夠充分;抗體特異性不好、抗體濃度太高等, 建議確定所選抗體是否合適、增加抗體稀釋 度、增加洗滌次數來改善。
Q38.陽性對照結果很漂亮,但是目標抗體沒有好的富集是為什么?
A38 :1)一般遇到這種情況,首先需要確認抗體質量本身,請選擇可靠的 ChIP 抗體,保證您的采購渠 道可信。
2)除非有足夠的證據,當使用自己免疫或定制的抗體時,請盡量不要選擇多肽免疫的抗體做 CUT&Tag 實驗。
3)不同轉錄因子與 DNA 結合強度不同,或者您的研究對象通過與其他蛋白形成復合物結合在特定的 DNA 片段上,必要時可以使用甲醛交聯。弱結合的轉錄因子容易從目標 DNA 上脫落,不完全是抗體 質量的問題。
4)由于轉錄因子在可以結合的目標片段往往遠少于組蛋白,PolII 等,打斷后的 DNA 量較少回收效率 低,建議適當增加細胞量。(由于 CUT&Tag 特異性很好,背景很低,研究轉錄因子時,電泳看不見 的 DNA 量足夠獲得很漂亮的結果,由于 DNA 量少,上機時按照傳統方法比較難通過測序公司質控, 需要與測序公司特別溝通。
Q39.CUT&Tag 重復性如何?如何評估 CUT&Tag 數據重復性?
A39 :1)基本分析方法參考 ChIP-Seq,同一組樣品做三組重復實驗,采用 deeptools 做 plotCorrelation 分析。
2)相關分析可以直接使用 BAM 比對文件,或分析 peak 文件,注意兩種不同的文件得到的相關性數值 會有較大差異。
3)做相關性分析時,采用泊松分布(pearson)會得到更好的分析靈敏度,注意使用泊松分布時應去除 特別多的 reads( (--removeOutliers )。
4)一般來說,同一次實驗相關性在 90%以上,不同條件的兩次實驗之間應大于 60%(見圖) Person 分 布,兩次實驗細胞數等有差別,BAM 文件組內相關性很好,組間60-70%;峰相關性很好。
Q40.與 ChIP-Seq 不同,由于 CUT&Tag 背景很低,不需要做input,數據分析時應注意哪些問題?
A40:CUT&Tag 貢獻信息的主要為單核小體與無核小體片段,因此建議序列比對時盡量選擇小片段如 小于 700bp , 在數據量足夠時可以只采用<350bp 的片段
[1] Sun S, Jiang Y, Zhang Q, et al. Znhit1 controls meiotic initiation in male germ cells by coordinating with Stra8 to activate meiotic gene expression. Dev Cell. 2022;57(7):901-913.e4. doi:10.1016/j.devcel.2022.03.006(IF:13.417)
[2] Yan QY, Lv JL, Shen XY, et al. Patchouli alcohol as a selective estrogen receptor β agonist ameliorates AD-like pathology of APP/PS1 model mice. Acta Pharmacol Sin. 2022;43(9):2226-2241. doi:10.1038/s41401-021-00857-4(IF:7.169)
[3] Liang T, Bai J, Zhou W, et al. HMCES modulates the transcriptional regulation of nodal/activin and BMP signaling in mESCs. Cell Rep. 2022;40(2):111038. doi:10.1016/j.celrep.2022.111038(IF:9.995)
[4] Jiang H, Bian W, Sui Y, et al. FBXO42 facilitates Notch signaling activation and global chromatin relaxation by promoting K63-linked polyubiquitination of RBPJ. Sci Adv. 2022;8(38):eabq4831. doi:10.1126/sciadv.abq4831(IF:14.957)
[5] Kong X, Yan K, Deng P, et al. LncRNA-Smad7 mediates cross-talk between Nodal/TGF-β and BMP signaling to regulate cell fate determination of pluripotent and multipotent cells. Nucleic Acids Res. 2022;50(18):10526-10543. doi:10.1093/nar/gkac780(IF:19.160)