lncRNA是一類長度大于200個核苷酸的RNA分子。根據lncRNA相對于附近編碼基因的位置將其分為antisense、intronic、intergenic、bidirectional和啟動子相關等5類lncRNA,這些lncRNA對編碼基因的調控方式大有不同。antisense lncRNA是一類與正向基因具有部分外顯子重疊的反義RNA,這種天然反義lncRNA的存在及位置的特殊性揭示其與正義鏈的作用往往比一般靶基因更密切。那么,antisense lncRNA作為數量較多的一類lncRNA,它是如何發揮作用的呢?下面從表觀遺傳學、轉錄及轉錄后水平進行簡單介紹。
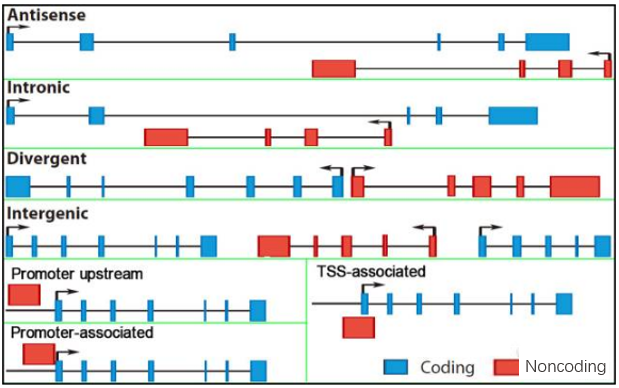
圖1 lncRNA相對于編碼基因位置
表觀遺傳調控
在不改變基因核苷酸序列的情況下,antisense lncRNA能夠結合DNA進行甲基化酶及染色質組蛋白修飾酶的招募,通過改變染色質狀態,調控正義基因的表達,并且產生可遺傳變異。像這種修飾可以小到作用于一個基因,大到影響整個染色體,比如Xist和反義Tsix二者介導的X染色體失活過程。
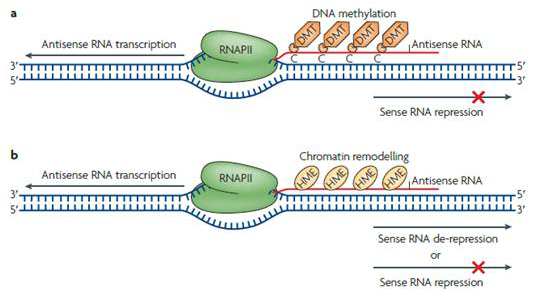
圖2 antisense lncRNA產生的表觀遺傳修飾
由于哺乳動物雌性含有兩條X染色體,而雄性只有一條X染色體,為了確保雄性(XY)和雌性(XX)的X基因劑量相等,雌性胚胎會發生單個X染色體永久失活,即“劑量補償”效應。非編碼RNA Xist是X染色體失活的主要參與基因,Xist僅在失活的X染色體中表達,其通過包裹合成它的X染色體,并結合Eed/PRC2復合物形成H3K27me3組蛋白修飾,進而抑制基因轉錄,致使X染色體失活。而在Xist基因3'端存在antisense lncRNA Tsix,Tsix的轉錄可以阻止Eed/PRC2募集到Xist啟動子區并且抑制H3K4甲基化,導致染色質結構發生改變,最終沉默Xist。
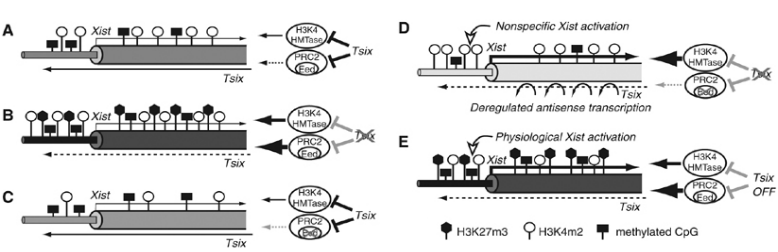
圖3 Tsix對Xist染色質的修飾狀態1
轉錄水平
由于antisense lncRNA和正義鏈基因方向相反,單單從空間位置來說,有模型表明當正/反義鏈同時轉錄形成RNA時,RNA聚合酶會相遇產生一定的空間阻礙,從而使RNA的轉錄受到抑制,像這種轉錄調控是由于基因本身方向相反導致,并不是由antisense lncRNA本身的調控功能引起對正義鏈基因轉錄的影響。
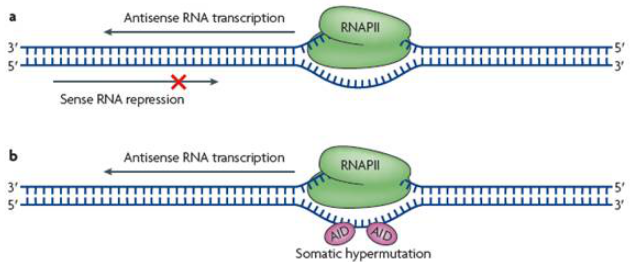
圖4 antisense lncRNA轉錄模式圖2
在細胞核內,antisense lncRNA能夠與正義鏈RNA通過堿基配對形成二聚體,二聚體的形成會導致部分外顯子的剪接位點被掩蓋掉,從而影響前體pre-mRNA的剪切過程,產生不同種類的mRNA成熟體。研究表明,antisense lncRNA除了通過二聚體形式調控pre-mRNA的成熟剪切,它也可以對核內mRNA的部分核苷酸進行替換編輯,如果蠅的反義Sas-10通過結合Rnp4f,將Rnp4f部分腺嘌呤替換為次黃嘌呤,從而降低Rnp4f mRNA的表達水平。
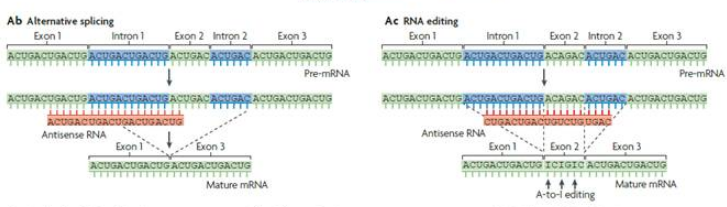
圖5 核內正/反義鏈RNA 堿基配對形成二聚體形式2
轉錄后水平
Antisense lncRNA不只在細胞核內影響正義基因的轉錄,正/反義鏈RNA二聚體同樣能夠在轉錄后水平調控基因的表達。antisense lncRNA可以結合正義基因mRNA 3’ UTR區,封閉大量miRNA的靶位點,阻礙了miRNA對靶基因的抑制作用,從而維持mRNA的穩定性。如反義lncRNA-PXN-AS1-L能夠結合PXN mRNA 3’ UTR,提高PXN mRNA的穩定性,最終促進癌基因RXN的表達和腫瘤細胞的生長3。
當然,antisense lncRNA同樣能夠吸附miRNA,進而減弱miRNA對靶基因的下調。另外,antisense lncRNA可與編碼蛋白基因的轉錄本形成互補雙鏈,在Dicer酶的作用下產生類似內源性的siRNA,從而沉默編碼基因的表達。
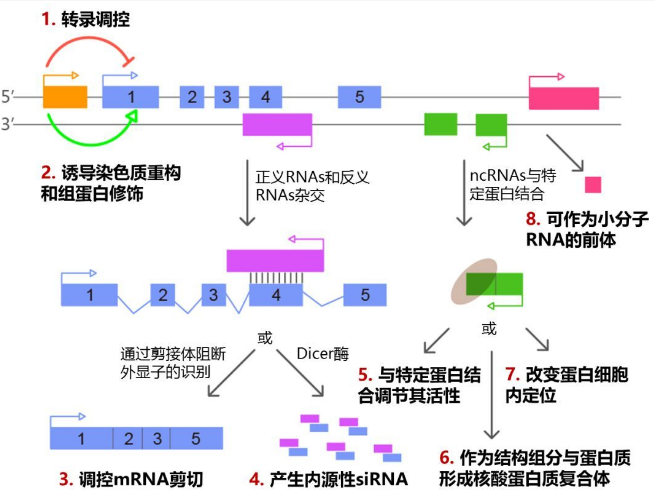
圖6 lncRNA作用機制
目前很多關于antisense lncRNA的調控機制不是很清楚,但很多研究表明其在機體的生長、發育和疾病發生過程中起著重要作用。所以,只要我們鍥而不舍深入挖掘,總會有機制明了的時候。
吉凱基因具有高效的質粒、病毒等工具表達載體,可以實現非編碼基因在細胞及動物水平上的過表達、敲低和敲除等目的,為非編碼RNA調控機制的研究提供便利,同時為廣大優秀的科研學者助力。

關注 | 病毒實驗幫
公眾號底部菜單欄【新功能】上線!
免費在線學習《國自然熱點研究》、《數據庫及軟件操作教程》
一鍵下載《病毒使用手冊》、《高分文獻》