
編者按
今天,我們特別關注一項于2021年2月發表在《Nature Communications》的新研究——《Genomic aberrations after short-term exposure to colibactin-producing E. coli transform primary colon epithelial cells》,該研究開發了來源于正常小鼠結腸上皮細胞的原代細胞培養模型,利用pks+大腸桿菌或大腸桿菌素合成缺陷的等基因突變體感染類器官和源自原代結腸上皮細胞的極化單層細胞,重現了感染的早期反應。
論文翻譯:何玲玲
01、研究背景
結直腸癌(CRC)是全球癌癥相關死亡的第二大原因。驅動癌變的基因突變已被很好地探索,例如Wnt、KRAS、TGF-β和p53通路[1]。這些通路中的突變組合負責上皮細胞的轉化[2,3],并且活躍的Wnt信號被認為是幾乎所有CRC的關鍵特征[4]。Wnt信號驅動結腸細胞增殖,僅在干細胞腔室中具有生理活性,而分化的細胞缺乏Wnt信號[5]。實驗和來自患者的數據表明,組成性活躍Wnt信號是CRC級聯的早期事件,通常通過APC突變和隨后的雜合性(LOH)丟失來實現[6]。最近的數據同樣表明,環境因素是CRC的重要驅動因素,結腸微生物群在CRC[7,8,9]的發生和進展中都起著重要作用,某些定殖于腸道的細菌似乎通過基因毒素直接影響上皮細胞的基因組完整性。
大腸桿菌素是由pks致病性島編碼的基因毒素,由幾種腸桿菌科成員產生,例如屬于系統發育類B2的大腸桿菌。pks+大腸桿菌感染真核細胞系可引起DNA雙鏈斷裂(DSBs),導致巨噬細胞增多和細胞周期阻滯[10]。在炎癥性腸病(IBD)、家族性腺瘤性息肉病(FAP)和CRC患者中,已經觀察到攜帶pks基因的結腸粘膜相關大腸桿菌的增加[11,12,13]。大腸桿菌素通過共價修飾使DNA烷基化,并在培養的哺乳動物細胞和體內形成穩定的腺嘌呤-大腸桿菌素復合物[14,15,16],從而在結直腸癌患者中留下特定的突變特征[17,18]。此外,在不同的FAP和CRC小鼠模型中,pks+大腸桿菌促進了腫瘤的發生[19]。值得注意的是,在這些小鼠模型中,腫瘤也可以在缺乏大腸桿菌素的情況下出現,這就不確定大腸桿菌素是否只是促進了腫瘤的發生,或者它是否也起到了致病作用。
目前為了研究pks+大腸桿菌在體外引起的基因組不穩定性(CIN),只對轉化或永生化的細胞系進行了感染,尚不清楚健康的原代結腸上皮細胞是否以同樣的方式對感染作出反應。為了解決這一問題,該研究團隊開發了來源于正常小鼠結腸上皮細胞的原代細胞培養模型。其利用pks+大腸桿菌或大腸桿菌素合成缺陷的等基因突變體感染類器官和源自原代結腸上皮細胞的極化單層細胞,重現了感染的早期反應。
02、主要研究成果
1、在原代結腸上皮細胞培養中,pks+大腸桿菌引起DNA損傷、巨噬細胞增多和多核細胞的形成
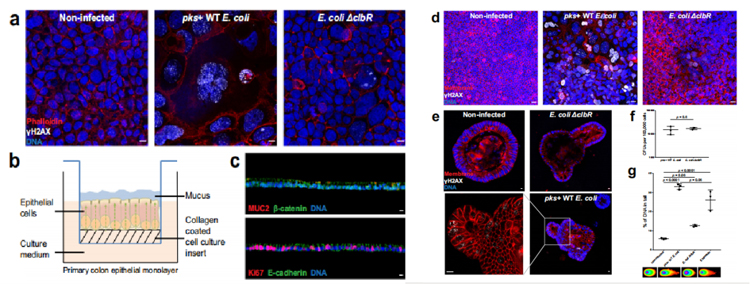
研究人員成功構建了在氣液界面中生長的極化上皮單層細胞(圖1b、1c),利用大腸桿菌素(pks+)野生型(WT) M1/5大腸桿菌感染該細胞3小時。為了確保大腸桿菌素具有特異性,研究人員使用大腸桿菌ΔclbR突變株作為陰性對照,該菌株對大腸桿菌素合成有缺陷。在MOI值為5的濃度下感染3小時下可檢測到γH2AX(DNA損傷修復蛋白)陽性細胞、巨噬細胞增多和多核細胞(圖1d),這些表型在感染ΔclbR突變體后未見,與Caco-2細胞系的表型相似(圖1a)。
研究人員進一步使用pks+大腸桿菌在MOI 5下感染小鼠結腸類器官3小時,類器官首先被分解,因此基部和根尖兩側都暴露在細菌中。3天后,在類器官重構后進行全載免疫熒光標記。同樣地,在感染pks+ WT大腸桿菌菌株后才觀察到γH2AX陽性細胞,這表明該系統適合研究大腸桿菌素對原代細胞的影響(圖1e)。
為了證實在原代細胞感染情況下觀察到的影響不是由于WT和ΔclbR突變型大腸桿菌之間的感染性差異,我們對類器官感染進行了集落形成單位(CFU)分析,證實了相同的定植率(圖1f)。為了進一步證實DSBs的存在,研究人員用中性彗星法測量了小鼠結腸類器官細胞中受損的DNA。結果顯示,與未感染的情況相比,pks+ WT大腸桿菌感染造成了更多的DNA損傷(圖1g)。
原文圖1
2、pks+大腸桿菌感染后類器官的wnt非依賴性生長
為進一步探索DNA修復機制下的非永生化原代細胞的反應,研究人員用pks+大腸桿菌感染初代極化上皮單層,免疫標記γ-H2AX和Ki67。感染3 h后,有部分細胞呈現γ-H2AX和Ki67雙陽性,表明它們在DNA損傷的情況下仍能增殖。這種不完全修復的DNA損傷下的細胞增殖可引起體細胞突變,這可能導致獨立于生態位因子;這種生態位逃逸是癌癥類器官的一個特征。
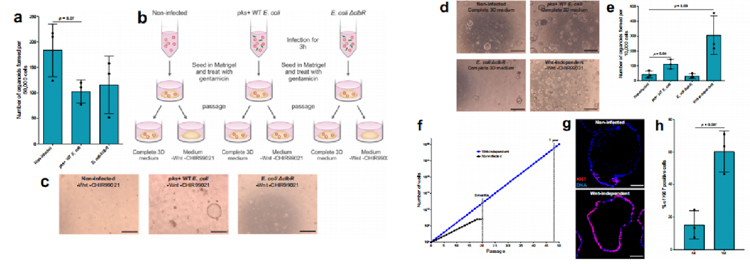
為了評估這類類器官形成能力,我們在MOI值為5的濃度下感染類器官3小時,將它們重新播種在Matrigel中,并用慶大霉素處理以殺死細菌。為了確定細胞死亡的程度,我們進行了一項類器官形成試驗,在每種條件下播種50,000個細胞。我們觀察到pks+ WT大腸桿菌感染的類器官形成效率降低了約40%,但WT和ΔclbR突變型大腸桿菌感染條件之間沒有差異(圖2a)。
由于CRC中最早和最常觀察到的突變是導致Wnt通路激活的突變,在接下來的傳代中,研究人員將類器官重新播種到含有Wnt和Wnt激動劑CHIR99021的完整3D培養基中,以及缺乏這兩種因子的培養基中(圖2b)。
結果顯示,未感染和ΔclbR突變感染的類器官在無wnt培養基中不能重新生長,而在pks+ WT大腸桿菌感染的條件下,一些類器官可重新形成(圖2c)。這些不依賴wnt的類器官可以進一步傳代和擴張。因此,將原代細胞短時間暴露于產生大腸桿菌素的大腸桿菌中,就足以引起體細胞適應,從而產生不依賴wnt的克隆。
原文圖2
3、不依賴wnt的類器官表現出增強的類器官形成能力、更高的增殖能力和壽命
為了表征感染對類器官生長的影響,研究人員測量了未感染、pks+ WT大腸桿菌感染和ΔclbR突變感染的類器官以及之前實驗中產生的不依賴wnt的類器官的類器官形成能力。結果顯示,與未感染或感染ΔclbR突變體的細胞相比,從WT pks+大腸桿菌感染條件下獲得的細胞具有更高的類器官形成能力。然而,來自wnt非依賴性類器官的細胞容量最高(圖2d, e),傳代率也更高,反映了它們的生長能力增加(圖2f)。因此,傳代7天后的類器官免疫標記顯示,與未感染的類器官相比,不依賴wnt的類器官具有更高比例的ki67陽性細胞(圖2g和h)。
4、不依賴Wnt的類器官顯示Wnt/β-catenin信號上調和分化基因下調
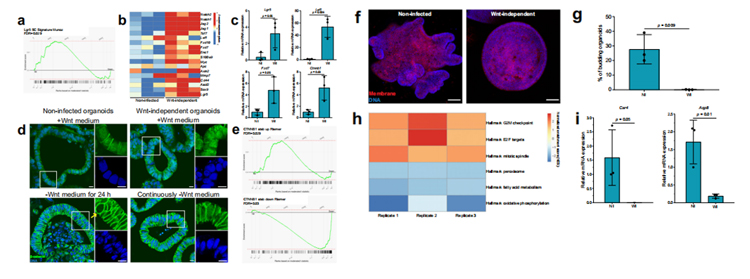
為了確定干細胞相關通路是否在不依賴wnt的類器官中上調研究人員對未感染和不依賴wnt的類器官進行了RNA測序及RT-qPCR。結果顯示,與未感染的類器官相比,不依賴wnt的類器官中Lgr5特征基因顯著富集(圖3a)、Wnt/β-catenin相關基因上調(圖3b、3c)。β-catenin免疫染色顯示,無論Wnt存在與否,核β-catenin都存在于不依賴wnt的類器官中,表明Wnt/β-catenin信號通路活躍。未感染的類器官在沒有Wnt的情況下保存24小時,在幾個細胞核中沒有細胞核β-catenin蛋白(圖3d)。此外,與未感染的類器官相比,與β-catenin的穩定性相關的Wnt信號基因組在不依賴wnt的類器官中分別顯示出顯著的正向和負向富集(圖3e),這表明不依賴wnt類器官對內源性Wnt具有更高的敏感性。
腸道類器官中Wnt信號的上調已被證明可導致細胞呈現增殖性祖細胞表型,從而導致細胞從不對稱的隱窩絨毛結構(未感染的類器官)轉變為球體、囊腫樣形態(不依賴wnt的類器官),缺乏分化的細胞類型(圖3f)。
此外,GSEA揭示了增殖相關的基因組上調、脂肪酸代謝相關的基因組下調,這些基因組在分化的結腸細胞中是活躍的(圖3h);腸細胞特異性分化基因碳酸酐酶4 (Car4)和水通道蛋白8 (Aqp8)在不依賴wnt的類器官中下調(圖3i)。因此,感染誘導的類器官wnt獨立性伴隨著增殖細胞表型和分化受損。
原文圖3
5、不依賴wnt的類器官具有主要的染色體畸變和高突變負荷
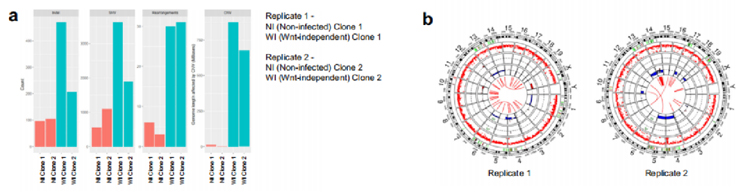
為了確定大腸桿菌素誘導的改變是否與突變有關,我們從未感染和不依賴wnt的類器官中產生克隆和混合培養物,并對兩個克隆中獲得的細胞進行全基因組(WGS)和全外顯子組測序(WXS)。結果顯示,與未感染的對照組相比,不依賴wnt類器官中存在更多的克隆(等位基因比例>25%)單核苷酸變異、插入和缺失突變,以及重排斷點。在由wnt非依賴性類器官產生Replicate 1和Replicate 2中,分別約有8.75億個和6.5億個堿基對受到拷貝數變異(CNV)的影響(圖4a)。因此,不依賴wnt的細胞表現出CNV的CIN特征,包括全染色體或染色體臂的主要損失(圖4b)。
原文圖4
03、編者點評
作為健康美麗產業CRO服務開拓者與引領者、斑馬魚生物技術的全球領導者,環特生物搭建了“斑馬魚、類器官、哺乳動物、人體”四位一體的綜合技術服務體系,開展健康美麗CRO服務、科研服務、智慧實驗室搭建三大業務。目前,環特類器官平臺已成功搭建多種腫瘤及正常類器官的培養平臺,歡迎有需要的讀者垂詢!
參考文獻
1. Fearon, E. R. & Vogelstein, B. A genetic model for colorectal tumorigenesis.Cell 61, 759–767 (1990).
2. Drost, J. et al. Sequential cancer mutations in cultured human intestinal stem cells. Nature 521, 43–47 (2015).
3. Matano, M. et al. Modeling colorectal cancer using CRISPR-Cas9-mediated engineering of human intestinal organoids. Nat. Med. 21, 256–262 (2015).
4. The Cancer Genome Atlas Network Comprehensive molecular characterization of human colon and rectal cancer. Nature 487, 330–337 (2012).
5. Barker, N. et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 449, 1003–1007 (2007).
6. Miyoshi, Y. et al. Somatic mutations of the APC gene in colorectal tumors:mutation cluster region in the APC gene. Hum. Mol. Genet. 1, 229–233 (1992).
7. West, N. R., McCuaig, S., Franchini, F. & Powrie, F. Emerging cytokine networks in colorectal cancer. Nat. Rev. Immunol. 15, 615–629 (2015).
8. Brentnall, T. A. et al. Mutations in the p53 gene: an early marker of neoplastic progression in ulcerative colitis. Gastroenterology 107, 369–378 (1994)
9. Fujita, M. et al. Genomic landscape of colitis-associated cancer indicates the impact of chronic inflammation and its stratification by mutations in the Wnt signaling. Oncotarget 9, 969–981 (2018).
10. Hussain, S. P. et al. Increased p53 mutation load in noncancerous colon tissue from ulcerative colitis: a cancer-prone chronic inflammatory disease. Cancer Res. 60, 3333–3337 (2000).