
編者按
急性心肌梗死(AMI)是一種危及生命的心血管急癥,以其高發(fā)病率、高死亡率成為常見的急危重癥疾病。與哺乳動(dòng)物不同,斑馬魚通過心肌細(xì)胞(CMs)的增殖來實(shí)現(xiàn)心臟的完全再生。
今天,我們分享2025年2月由德國(guó)烏爾姆大學(xué)、博羅尼亞大學(xué)等研究團(tuán)隊(duì)聯(lián)合在《Nature Communications》雜志上發(fā)表的最新研究,該研究表明克服復(fù)制壓力的能力是增強(qiáng)斑馬魚心臟再生能力的關(guān)鍵因素,并揭示了BMP信號(hào)在促進(jìn)無壓力DNA復(fù)制中的作用。
研究發(fā)現(xiàn),再生的心肌細(xì)胞經(jīng)歷DNA復(fù)制壓力,是哺乳動(dòng)物衰老過程中組織再生能力下降的原因之一。通過抑制ATM和ATR激酶,表明DNA損傷應(yīng)答信號(hào)對(duì)斑馬魚心臟再生至關(guān)重要。利用轉(zhuǎn)基因和突變體操縱骨形態(tài)發(fā)生蛋白(bone morphogenetic protein,BMP)-Smad信號(hào)表明,BMP信號(hào)可緩解心肌細(xì)胞的復(fù)制壓力,也能使新生小鼠的心肌細(xì)胞、人類成纖維細(xì)胞、人造血干細(xì)胞(HSCs)及祖細(xì)胞(HSPCs)免受復(fù)制壓力的影響。DNA纖維測(cè)定技術(shù)分析表明,BMP信號(hào)可以在復(fù)制壓力導(dǎo)致的停滯后重新啟動(dòng)復(fù)制叉。
文章題目
BMP signaling promotes zebrafish heart regeneration via alleviation of replication stress
雜志:Nature Communications(IF=16.6)
發(fā)表時(shí)間:2025年2月17日
作者:Mohankrishna Dalvoy Vasudevarao, Hartmut Geiger &Gilbert Weidinger et al.
單位:德國(guó)烏爾姆大學(xué)、博羅尼亞大學(xué)等
DOI: 10.1038/s41467-025-56993-6
01、研究亮點(diǎn)
• 首次證明斑馬魚心臟再生過程中,增殖的心肌細(xì)胞(CMs)會(huì)經(jīng)歷顯著的DNA復(fù)制壓力,這被認(rèn)為是哺乳動(dòng)物衰老過程中組織再生能力下降的關(guān)鍵限制因素;
• 發(fā)現(xiàn)骨形態(tài)發(fā)生蛋白(BMP)信號(hào)通路,特別是BMP7a配體通過Smad依賴性途徑,在緩解心肌細(xì)胞復(fù)制壓力中的核心作用。BMP信號(hào)通過促進(jìn)停滯的復(fù)制叉重新啟動(dòng)(fork re-start)來實(shí)現(xiàn)這一功能;
• 證明DNA損傷響應(yīng)通路對(duì)斑馬魚心臟再生至關(guān)重要,抑制ATM及ATR激酶會(huì)阻斷心肌細(xì)胞增殖和再生;
• 研究發(fā)現(xiàn),斑馬魚心臟再生并非避免了復(fù)制壓力,而是具備了BMP信號(hào)等強(qiáng)大的機(jī)制來克服這種壓力,為理解再生能力差異和開發(fā)抗衰老、促再生策略提供了新視角。
02、研究結(jié)果
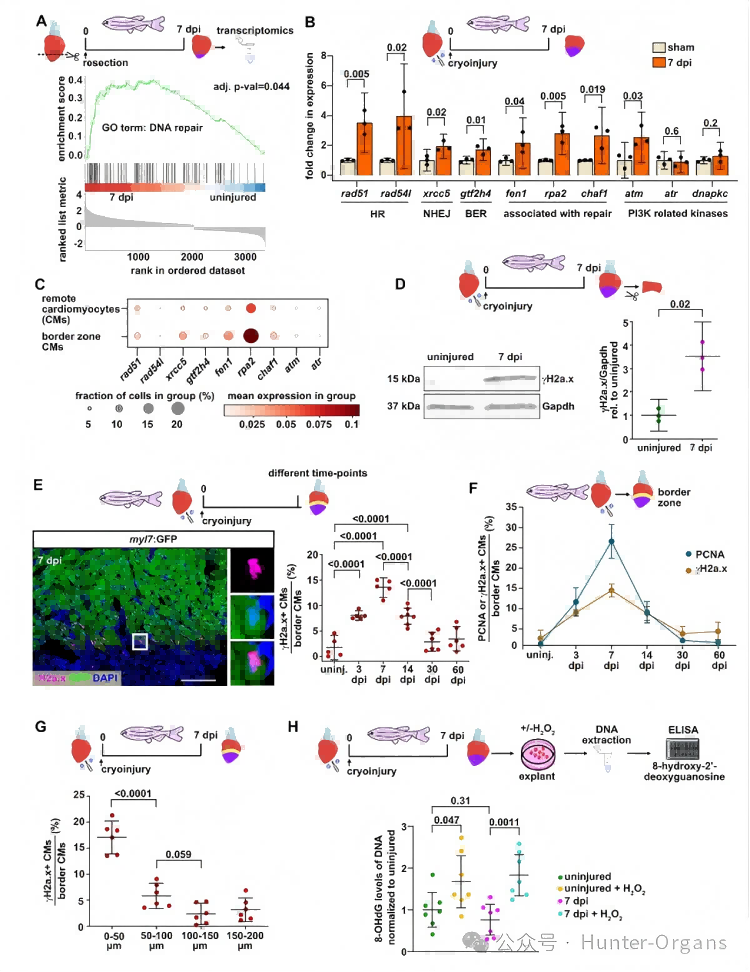
1. 斑馬魚心臟再生過程中心肌細(xì)胞變?yōu)?gamma;H2a.x陽性
為了探究心臟再生機(jī)制,研究人員分析了斑馬魚心臟轉(zhuǎn)錄組,注意到與DNA修復(fù)相關(guān)的基因特征在損傷后7天(dpi)富集,這代表了CM增殖的高峰期。qRT-PCR顯示,在冷凍損傷的心臟中,多種DNA損傷反應(yīng)通路的基因在7dpi時(shí)上調(diào)。位于傷口邊緣的心肌細(xì)胞(CMs)是心臟再生的關(guān)鍵,用于分化、增殖以替換受損的心肌。利用單細(xì)胞測(cè)序發(fā)現(xiàn),一些上調(diào)的DNA修復(fù)相關(guān)基因在7dpi時(shí),其傷口邊緣區(qū)的心肌細(xì)胞(CMs)表達(dá)比遠(yuǎn)端的更強(qiáng)。
通過蛋白質(zhì)印跡檢測(cè)到,7dpi時(shí)心臟中的DNA損傷標(biāo)志物——γH2a.x水平升高;免疫熒光顯示,在7dpi時(shí),傷口邊緣的一部分心肌細(xì)胞呈γH2a.x陽性,且與細(xì)胞周期的進(jìn)展密切相關(guān)。γH2a.x陽性心肌細(xì)胞的數(shù)量隨傷口距離而減少。經(jīng)心尖切除術(shù)(AR)的再生新生心臟組織(AR-Neo-EVs)中也觀察到了γH2a.x陽性心肌細(xì)胞。這些數(shù)據(jù)表明,再生心臟中的心肌細(xì)胞經(jīng)歷了DNA損傷,但與損傷類型無關(guān),且不太可能直接由損傷本身引起。
隨后,研究人員利用DNA氧化損傷的重要標(biāo)志物——8-羥基脫氧鳥苷(8-OHdG)進(jìn)行了酶聯(lián)免疫吸附試驗(yàn) (ELISA) 。實(shí)驗(yàn)結(jié)果顯示,心臟外植體的過氧化物處理誘導(dǎo)了DNA氧化,但與未受傷的心臟相比,僅冷凍損傷并未增加8-OHdG水平。這表明,斑馬魚的DNA氧化損傷不太可能是心肌細(xì)胞中γH2a.x陽性(即DNA損傷反應(yīng))的原因。
2. 心肌細(xì)胞在再生過程中經(jīng)歷復(fù)制壓力
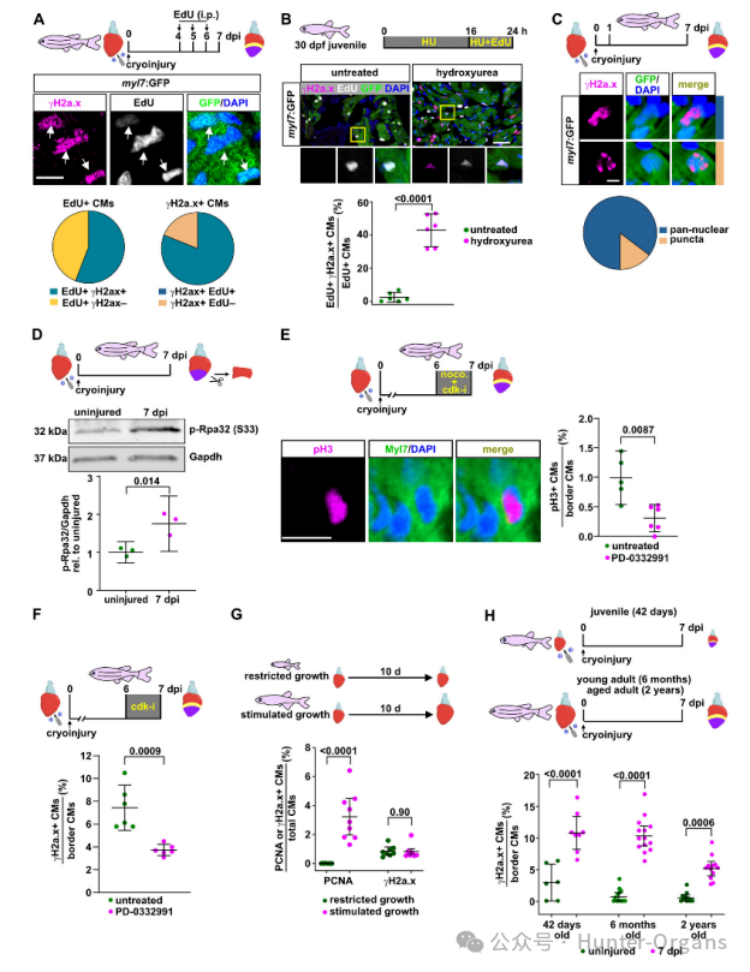
γH2a.x積累的時(shí)間和空間分布特征表明,在傷口邊緣進(jìn)入細(xì)胞周期的心肌細(xì)胞經(jīng)歷了DNA損傷。研究人員在7dpi時(shí)對(duì)γH2a.x和EdU標(biāo)記進(jìn)行染色后發(fā)現(xiàn),γH2a.x陽性主要出現(xiàn)在最近或仍在分裂的心肌細(xì)胞,這表明心肌細(xì)胞變?yōu)?gamma;H2a.x陽性是因?yàn)樵谠偕^程中其進(jìn)入細(xì)胞周期時(shí)經(jīng)歷了復(fù)制壓力。
在未受傷心臟中,只有0.62%的心肌細(xì)胞是γH2a.x陽性,而用羥基脲(HU)處理,其消耗細(xì)胞的核苷酸從而引起復(fù)制壓力,誘導(dǎo)了γH2a.x陽性。EdU和γH2a.x的雙重染色顯示,在分裂中的心肌細(xì)胞,γH2a.x積累很少見(2.3%),但HU處理可顯著增加其比例。同樣,在冷凍損傷的心臟中,HU處理增加了γH2a.x陽性心肌細(xì)胞的比例。這些數(shù)據(jù)表明,γH2a.x通常并不標(biāo)記分裂中的斑馬魚心肌細(xì)胞,而是作為復(fù)制壓力的可靠指標(biāo)。
γH2a.x在細(xì)胞核中的定位也指示了DNA損傷的不同類型。如,電離輻射主要導(dǎo)致雙鏈斷裂,從而引起再生心臟的心肌組織中出現(xiàn)明顯的點(diǎn)狀γH2a.x積累,而HU則引起泛核γH2a.x染色。在未受干擾的再生心臟中,84%的γH2a.x陽性心肌細(xì)胞顯示泛核γH2a.x分布。磷酸化Rpa32是形成單鏈DNA結(jié)合蛋白R(shí)PA復(fù)合物的一部分,磷酸化Rpa32積累被認(rèn)為是復(fù)制壓力的特異性指標(biāo)。蛋白質(zhì)印跡顯示,在7dpi時(shí),心肌心室組織中p-Rpa32(S33)水平顯著增加。因此,通過γH2a.x積累可以發(fā)現(xiàn),傷口邊緣的心肌細(xì)胞經(jīng)歷復(fù)制壓力。
為了進(jìn)一步研究再生心臟中心肌細(xì)胞增殖與γH2a.x積累之間的相關(guān)性,研究人員使用CDK4/6抑制劑PD-0332991來直接抑制細(xì)胞周期進(jìn)程,結(jié)果發(fā)現(xiàn),7dpi時(shí)顯著減少分裂期pH3陽性心肌細(xì)胞和γH2a.x陽性心肌細(xì)胞的數(shù)量。需要注意的是,生理狀態(tài)下心肌細(xì)胞的增殖不會(huì)引起γH2A.X陽性的累積,而在心臟受傷時(shí),γH2A.X會(huì)在心肌細(xì)胞中累積。總之,這些發(fā)現(xiàn)表明,心肌細(xì)胞的復(fù)制壓力是由再生需求誘導(dǎo)的特異性引起,而不是由生理性生長(zhǎng)誘導(dǎo)的。
此外,通過HU處理誘導(dǎo)的外源性復(fù)制壓力并不影響心肌細(xì)胞去分化的三個(gè)指標(biāo),即祖細(xì)胞標(biāo)志物gata4調(diào)控區(qū)活性的轉(zhuǎn)基因的上調(diào)、胚胎肌球蛋白(embMHC)的上調(diào)或肌節(jié)的解體,這表明心肌細(xì)胞去分化獨(dú)立發(fā)生。
3. 心肌細(xì)胞復(fù)制壓力可能并非由DNA損傷積累或轉(zhuǎn)錄碰撞引起
在幼魚期(42天)、成年早中期(6個(gè)月)和老年期(2歲)的斑馬魚再生心臟中,并未觀察到γH2a.x陽性心肌細(xì)胞比例的增加,這表明心肌細(xì)胞復(fù)制壓力不太可能由存在的DNA損傷阻礙復(fù)制叉引起。而針對(duì)活性、延伸的磷酸化RNA聚合酶II的免疫染色顯示,傷口邊緣心肌細(xì)胞的總體轉(zhuǎn)錄水平低于心室其他區(qū)域,這表明心肌細(xì)胞的增殖也不大可能是由轉(zhuǎn)錄碰撞引起的。
4. DNA損傷信號(hào)傳導(dǎo)對(duì)心臟再生是必需的
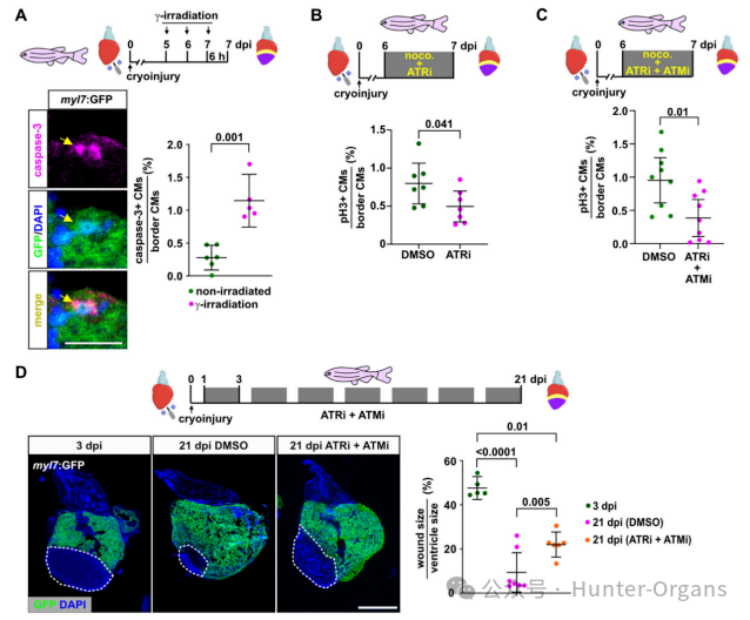
隨后,研究人員探究了經(jīng)歷復(fù)制壓力的心肌細(xì)胞是否會(huì)衰老或凋亡。在7dpi時(shí),衰老標(biāo)志物β-半乳糖苷酶在心外膜細(xì)胞中表達(dá)上調(diào),而非心肌細(xì)胞中,這與先前在再生斑馬魚和新生小鼠心臟中的報(bào)道一致。用HU處理則并未誘導(dǎo)心肌細(xì)胞衰老,因此,經(jīng)歷復(fù)制壓力的心肌細(xì)胞不會(huì)衰老。
在未受干擾的再生心臟中,通過檢測(cè)Caspase3的積累發(fā)現(xiàn),只有0.27%的傷口邊緣心肌細(xì)胞凋亡,而電離輻射可以誘導(dǎo)了心肌細(xì)胞凋亡。這表明,盡管斑馬魚心肌細(xì)胞保留了響應(yīng)DNA損傷誘導(dǎo)促凋亡信號(hào)的能力,但經(jīng)歷復(fù)制壓力的斑馬魚心肌細(xì)胞能克服復(fù)制壓力并繼續(xù)增殖。
基于此,感知復(fù)制壓力的DNA損傷通路可能對(duì)心臟再生是必需的。因此,研究人員又利用ATM和ATR激酶的抑制劑來分別介導(dǎo)對(duì)DNA損傷和復(fù)制壓力的細(xì)胞反應(yīng),并觀察其在斑馬魚中是否有效。研究發(fā)現(xiàn),ATM抑制劑KU55933和ATR抑制劑VE821是無毒的,其不影響胚胎發(fā)育,但用ATR抑制劑處理的胚胎在單獨(dú)使用小劑量的HU中出現(xiàn)壞死,而ATM抑制劑則加劇了低劑量電離輻射的影響,表明這些抑制劑阻斷了斑馬魚細(xì)胞響應(yīng)復(fù)制壓力,以及其他形式的DNA損傷激活修復(fù)通路的能力。
研究人員還發(fā)現(xiàn)用ATR抑制劑VE821處理斑馬魚24小時(shí)后,在7dpi時(shí)減少再生心臟中的心肌細(xì)胞有絲分裂。已知ATM和ATR激酶通路可以交叉作用,以促進(jìn)復(fù)制壓力反應(yīng),聯(lián)合使用ATM和ATR抑制劑比單獨(dú)使用ATR抑制劑,更顯著地抑制了心肌細(xì)胞的有絲分裂,且兩種抑制劑聯(lián)合治療也損害了心臟的再生修復(fù)。總之,這表明,由ATR介導(dǎo),并有ATM參與的DNA損傷修復(fù)通路的激活,對(duì)再生性心肌細(xì)胞增殖和心臟再生至關(guān)重要。
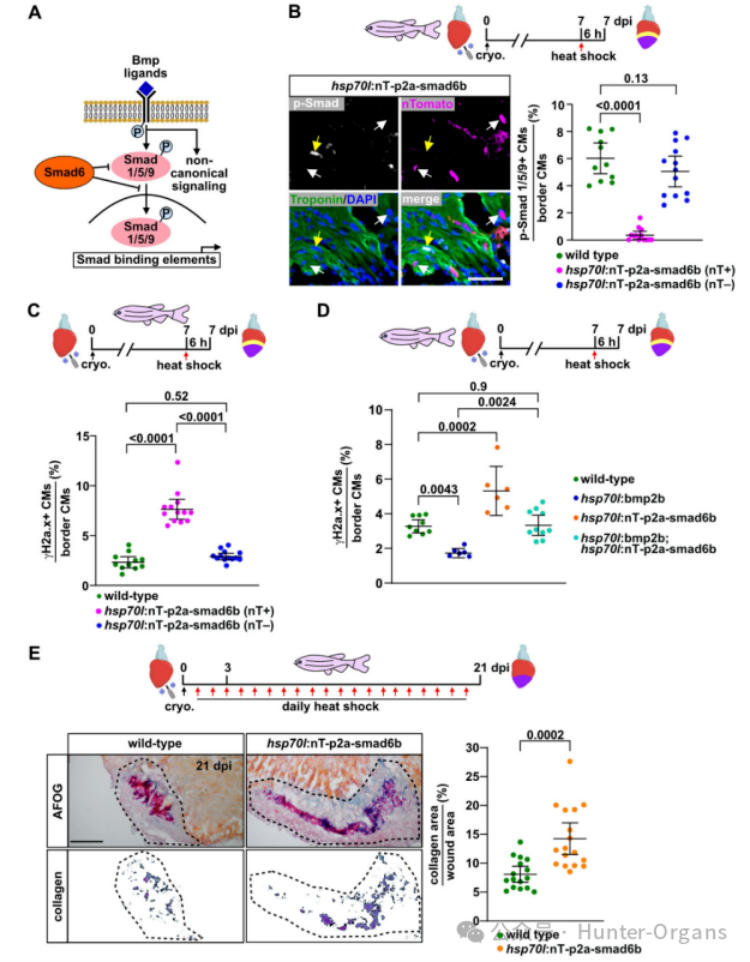
5. BMP信號(hào)傳導(dǎo)緩解心肌細(xì)胞復(fù)制壓力
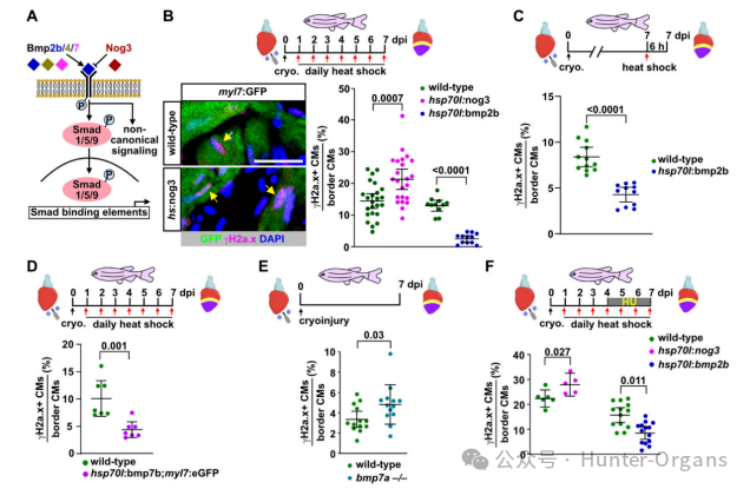
我們先前已證明在斑馬魚心臟再生過程中,BMP信號(hào)在傷口邊緣的心肌細(xì)胞中CMs中被激活,但在生理性生長(zhǎng)條件下則不會(huì)被激活,且它對(duì)再生性心肌細(xì)胞的增殖至關(guān)重要。因此,研究人員探究了BMP信號(hào)是否調(diào)節(jié)斑馬魚心肌細(xì)胞的復(fù)制壓力,結(jié)果顯示,表達(dá)nog3的斑馬魚在7dpi時(shí)顯示出更高比例的γH2a.x陽性心肌細(xì)胞,而過表達(dá)BMP配體bmp2b則有效地減少了γH2a.x陽性心肌細(xì)胞的數(shù)量。在7dpi時(shí),單次誘導(dǎo)bmp2b過表達(dá)可以在6小時(shí)內(nèi)顯著減少γH2a.x陽性心肌細(xì)胞的數(shù)量,這表明BMP信號(hào)可以直接作用于經(jīng)歷復(fù)制壓力的增殖心肌細(xì)胞。
過表達(dá)bmp7b或bmp4也可以減少γH2a.x陽性心肌細(xì)胞的比例。在7dpi時(shí),γH2a.x陽性心肌細(xì)胞的比例增加,p-Smad1/5/9水平降低,而突變體和野生型心臟之間則沒有形態(tài)學(xué)差異。因此,在再生過程中,內(nèi)源性BMP信號(hào)緩解心肌細(xì)胞復(fù)制壓力是必需的,且Bmp7a作為不可替代的配體也在其中發(fā)揮作用。有趣的是,在HU處理的斑馬魚中,nog3過表達(dá)進(jìn)一步增加了γH2a.x陽性心肌細(xì)胞的比例,而bmp2b過表達(dá)則顯著降低了這一比例。這些結(jié)果表明,BMP信號(hào)不僅可以緩解內(nèi)源性復(fù)制壓力,也能緩解外源性復(fù)制壓力。
6. BMP通過Smad發(fā)揮作用以抑制復(fù)制壓力
BMP配體可以激活Smad依賴性和非依賴性信號(hào)通路。為了探究BMP是否通過Smad緩解斑馬魚心臟的復(fù)制壓力,研究人員創(chuàng)建了轉(zhuǎn)基因品系hsp70l:nT-p2a-smad6bulm16Tg來比較nT+和nT-的心肌細(xì)胞。在7dpi熱休克后6小時(shí)內(nèi),nT+心肌細(xì)胞通過磷酸化Smad1/5/9核積累顯示出Smad信號(hào)傳導(dǎo)被抑制,nT+心肌細(xì)胞比野生型或nT-心肌細(xì)胞呈γH2a.x陽性的頻率高出3倍。因此,BMP/Smad信號(hào)傳導(dǎo)在心肌細(xì)胞中可以自主緩解復(fù)制壓力。
為了驗(yàn)證過表達(dá)的BMP配體是否僅通過Smad通路發(fā)揮作用,研究人員分析后發(fā)現(xiàn),bmp2b過表達(dá)增加了p-Smad1/5/9+心肌細(xì)胞的比例,而smad6b降低了該比例,但在雙轉(zhuǎn)基因魚中,p-Smad1/5/9+心肌細(xì)胞的比例與野生型中觀察到的一致。僅表達(dá)bmp2b的斑馬魚顯示γH2a.x+心肌細(xì)胞數(shù)量顯著減少,但smad6b的過表達(dá)可以逆轉(zhuǎn)bmp2b過表達(dá)對(duì)γH2a.x的影響。總之,這些數(shù)據(jù)表明BMP信號(hào)直接在心肌細(xì)胞中,通過Smad發(fā)揮作用,以緩解復(fù)制壓力。
為了探究抑制BMP/Smad信號(hào)傳導(dǎo)對(duì)形態(tài)學(xué)心臟再生和瘢痕形成的長(zhǎng)期影響,研究人員對(duì)冷凍損傷的轉(zhuǎn)基因斑馬魚及其野生型斑馬魚每日熱休克,連續(xù)處理21天,評(píng)估傷口大小和膠原沉積。在3dpi時(shí)傷口大小沒有差異,但冷凍損傷心臟在21 dpi時(shí)顯示出更大的傷口,且傷口含有許多膠原。這表明,BMP/Smad信號(hào)傳導(dǎo)對(duì)心臟再生和瘢痕消退是必需的。
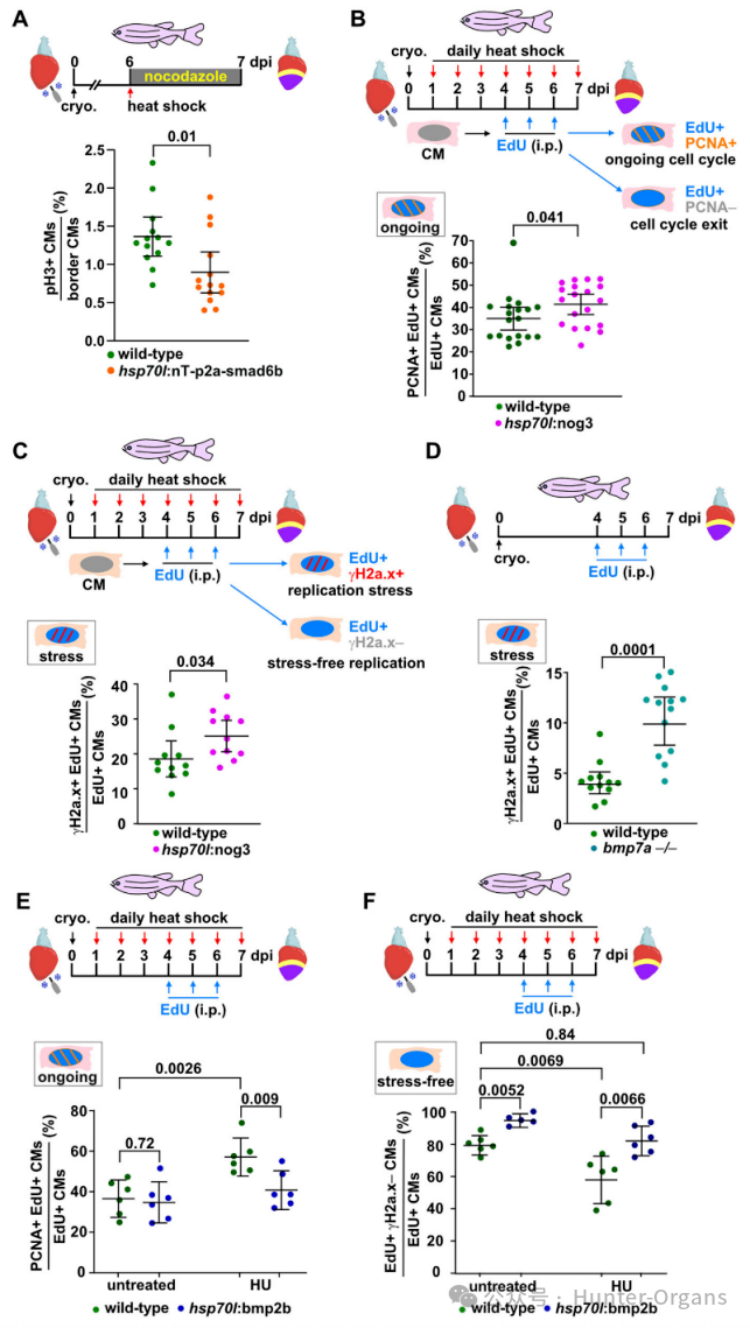
7. BMP信號(hào)傳導(dǎo)促進(jìn)無壓力復(fù)制和有絲分裂進(jìn)程
在冷凍損傷的斑馬魚中,單次誘導(dǎo)smad6b表達(dá)在6小時(shí)內(nèi)減少了pH3陽性有絲分裂心肌細(xì)胞的比例。通過重復(fù)EdU注射標(biāo)記受傷的野生型和轉(zhuǎn)基因斑馬魚,觀察其增殖的心肌細(xì)胞3天,并在7dpi時(shí)染色,發(fā)現(xiàn)單個(gè)心肌細(xì)胞不會(huì)經(jīng)歷多輪細(xì)胞分裂,在4、5或6dpi時(shí)增殖的少數(shù)心肌細(xì)胞在7dpi時(shí)仍在增殖,此時(shí)許多心肌細(xì)胞已不再增殖。在熱休克的野生型斑馬魚中,在7dpi時(shí)只有約35%的EdU+心肌細(xì)胞是PCNA+的,通過過表達(dá)nog3抑制BMP信號(hào)傳導(dǎo)增加了EdU+PCNA+心肌細(xì)胞的比例,smad6b過表達(dá)增加了PCNA+ CMs的比例,但減少了CM有絲分裂,這表明smad6b誘導(dǎo)了心肌細(xì)胞周期延遲,導(dǎo)致正在進(jìn)行細(xì)胞周期的心肌細(xì)胞增加。
隨后,研究人員又重復(fù)EdU注射標(biāo)記增殖的心肌細(xì)胞,對(duì)EdU和γH2a.x進(jìn)行染色,發(fā)現(xiàn)EdU+γH2a.x陽性心肌細(xì)胞經(jīng)歷復(fù)制壓力,而僅EdU+的心肌細(xì)胞則無壓力復(fù)制。過表達(dá)nog3會(huì)增加EdU+γH2a.x陽性心肌細(xì)胞的。總的來說,這表明,BMP信號(hào)傳導(dǎo)特異性促進(jìn)心肌細(xì)胞增殖,使心肌細(xì)胞能夠克服復(fù)制壓力,并進(jìn)入有絲分裂。
接下來,為了探究BMP信號(hào)激活能否促進(jìn)心肌細(xì)胞細(xì)胞周期進(jìn)程和無壓力復(fù)制,研究人員發(fā)現(xiàn),在未受干擾的條件下,bmp2b過表達(dá)不會(huì)降低PCNA+ EdU+ 心肌細(xì)胞的比例,但能緩解由HU處理引起的數(shù)量增加。無論是在未受干擾條件下還是在HU誘導(dǎo)的復(fù)制壓力下,bmp2b過表達(dá)都能增加經(jīng)歷“無壓力”復(fù)制的EdU+ γH2a.x-心肌細(xì)胞的比例。
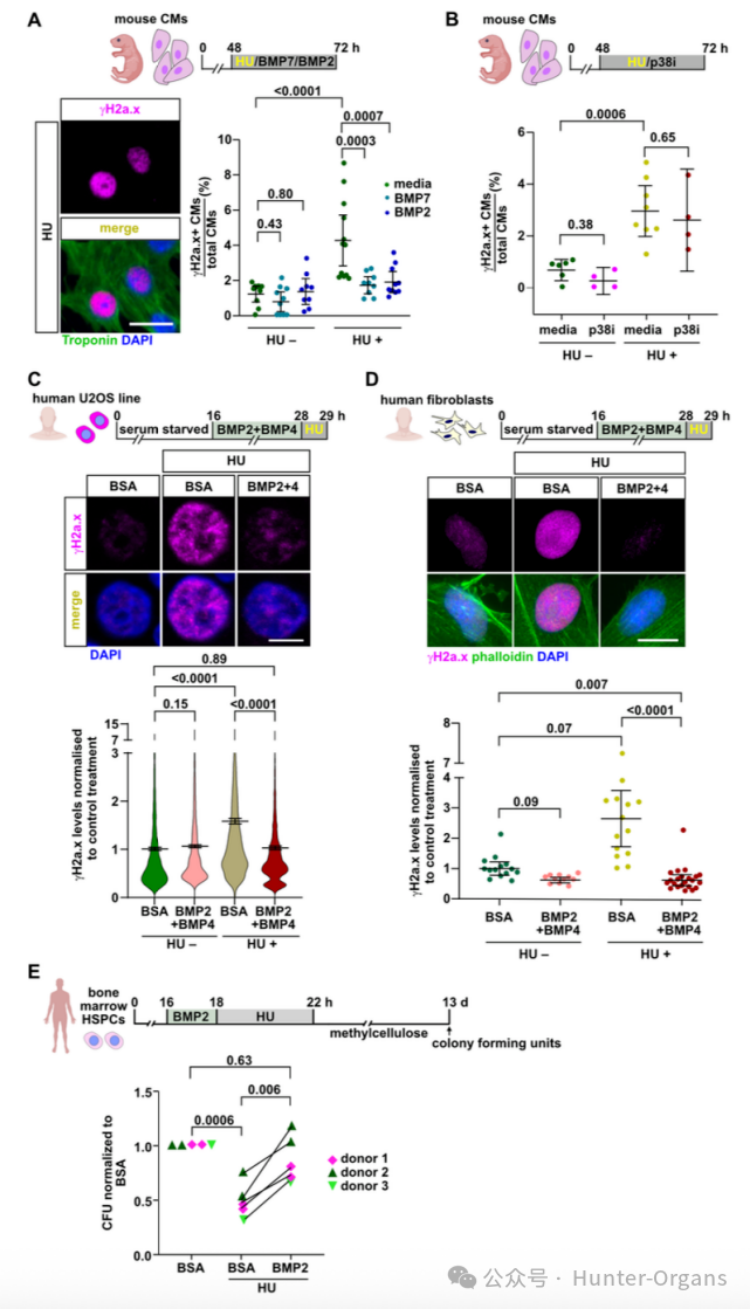
8. BMP信號(hào)在哺乳動(dòng)物及人類細(xì)胞中的保守性
隨后,研究人員進(jìn)一步探究了BMP信號(hào)在哺乳動(dòng)物及人類細(xì)胞中緩解復(fù)制壓力的能力。哺乳動(dòng)物實(shí)驗(yàn)結(jié)果表明,成纖維細(xì)胞/U2OS細(xì)胞中,BMP2+BMP4預(yù)處理可防止HU誘導(dǎo)的γH2a.x積累;造血干細(xì)胞/祖細(xì)胞 (HSPCs)中,BMP2預(yù)處理可完全挽救HU對(duì)原代人骨髓HSPCs集落形成能力的損傷;重組BMP2或BMP7蛋白處理可保護(hù)培養(yǎng)的新生小鼠心肌細(xì)胞免受HU誘導(dǎo)的復(fù)制壓力,減少γH2a.x+細(xì)胞,且這種保護(hù)作用具有特異性。
通過DNA纖維鋪展分析顯示,BMP信號(hào)在人臍帶血HSPCs/U2OS細(xì)胞中表現(xiàn)為CldU/IdU軌道長(zhǎng)度增長(zhǎng), BMP2或BMP4處理能增加復(fù)制叉的進(jìn)程速度;在U2OS細(xì)胞中,HU誘導(dǎo)復(fù)制叉停滯的模型則能通過BMP2或BMP4預(yù)處理,顯著提高復(fù)制叉再啟動(dòng)的比例。BMP信號(hào)能增強(qiáng)復(fù)制動(dòng)力學(xué),少部分通過增強(qiáng)復(fù)制叉重新啟動(dòng)能力來緩解復(fù)制壓力。
03、編者點(diǎn)評(píng)
本研究利用斑馬魚揭示了復(fù)制壓力是限制組織再生,尤其是心臟再生的關(guān)鍵瓶頸,并發(fā)現(xiàn)BMP-Smad信號(hào)通路作為克服這一瓶頸的核心保守機(jī)制,為理解再生生物學(xué)和開發(fā)再生醫(yī)學(xué)策略提供了重要的新見解。
作為健康美麗產(chǎn)業(yè)CRO服務(wù)開拓者與引領(lǐng)者、斑馬魚生物技術(shù)的全球領(lǐng)導(dǎo)者,環(huán)特生物搭建了“斑馬魚、類器官、哺乳動(dòng)物、人體”多維生物技術(shù)服務(wù)體系,開展健康美麗CRO服務(wù)、科研服務(wù)、智慧實(shí)驗(yàn)室搭建三大業(yè)務(wù)。目前,環(huán)特已建立200多種斑馬魚模型,胃癌、腦類器官、心臟類器官及各種腫瘤類器官培養(yǎng)平臺(tái),歡迎有需要的讀者垂詢!