?CRISPR/Cas9作為一項有力的基因編輯工具,已經被廣泛應用于生物醫學研究領域。隨著研究的深入,越來越多與其相關的研究工具已經被開發出來,包括基因敲除,基因過表達,將外源片段定點整合進入基因組,單堿基編輯,文庫高通量篩選等。
目前已有不少在體的研究也都在利用CRISPR/Cas9技術。在結合AAV病毒載體后,擁有廣泛的應用前景和臨床意義1。但很多在體研究還局限在敲除基因上面,原因是CRISPR/Cas9介導的KO依賴的是DNA雙鏈斷裂修復后引入或者缺失片段導致的移碼,相對較容易實現。如果想要精確的編輯某個基因,例如進行單堿基突變或者插入某個片段或者精確刪除某個DN**段則難度較高。
因為常規的看法是,這類精確編輯依賴的是HDR(同源重組介導的修復),但HDR的編輯效率非常低,體外研究通常占比低于5%。且HDR修復通路具有強烈的細胞周期依賴,發生在S/G2期,所以利用HDR來進行在體的基因編輯其效果受到很大限制,應用較少。
特在此為大家推薦其它一些高效且精確的基因編輯策略或者技術,這些方案都可以通過AAV或者慢病毒載體來實現。
# 單堿基編輯技術
在單堿基編輯技術出現之前,為了在基因上引入單堿基突變,只能依賴HDR修復途徑。而這個過程需要產生DNA雙鏈斷裂,還要引入donor序列,且效率非常低。
將胞嘧啶脫氨酶rAPOBEC1融合到dCas9的N端使得該融合蛋白可以將胞嘧啶(C)的氨基水解掉,從而變成尿嘧啶(U),這個水解過程發生在與sgRNA非互補的鏈上。由于尿嘧啶不是DNA的常規組分,細胞內的堿基切除修復通路(BER),將會識別DNA中的尿嘧啶,并將其切除,而這將抑制單堿基編輯的效率。一段尿嘧啶糖基化酶抑制劑(UGI)序列被融合到dCas9蛋白的C端,可以有效地阻止編輯點附近尿嘧啶被切除。
此時該位點的序列為U:G配對,隨著著細胞分裂將產生兩個子細胞,在該位點的序列分別為U:A和C:G。其中U:A的子細胞序列進一步分裂或者通過BER修復就變成T:A配對,最終完成C:G到T:A的轉變2。
由于最后一步的編輯依賴細胞分裂,為了降低這種依賴性,并進一步提高編輯效率,如何將非分裂細胞中的U:G突變成U:A就成了下一個研究的關鍵點。細胞內存在另外一條錯配修復通路(MMR),可以將錯配的堿基進行修復,例如前面不匹配的U:G。
如何讓MMR將U:G配對修復為U:A而非C:G呢?MMR會將含有缺刻的一條鏈識別為新合成的DNA鏈,從而切除這條鏈上錯配的堿基,并以另外一條鏈為模板進行修復。因此通過將dCas9的HNH酶切位點恢復活性,就可以切割與sgRNA互補的DNA鏈,即U:G配對中堿基G所在的鏈(圖1)。
相對于利用HDR引入點突變,單堿基編輯的效率明顯提高,達到15%-75%。這意味著如果在體實驗需要引入或者修正點突變,完全可以通過單堿基編輯技術實現。前面介紹的這個技術可以實現CT或者GA的突變,而如果將這個系統中胞嘧啶脫氨酶替換成腺嘌呤(A)脫氨酶,則可以實現AG或者TC的突變3。并且更適合AAV載體的saCas9也開發出了相關的系統4。
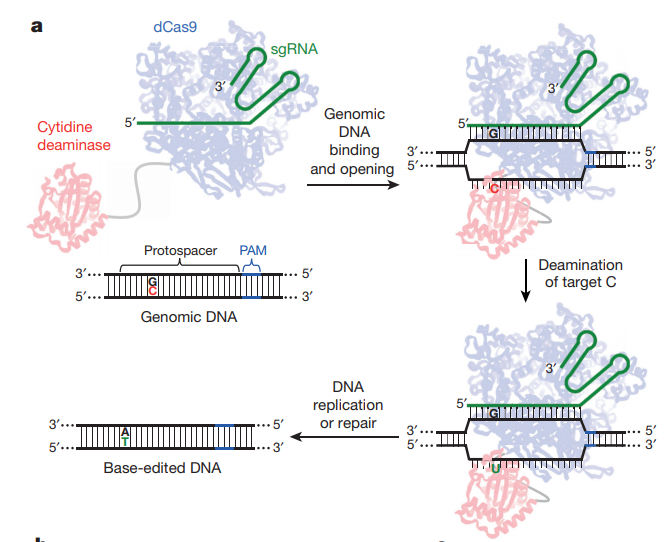
圖 1 單堿基編輯示意圖
#非同源依賴的片段整合(HITI)
如果想在體地在基因中引入某些片段,HDR的低效同樣使人望而卻步。在此也為大家介紹一項更高效的片段整合的技術HITI。眾所周知,末端連接修復通路(NHEJ或者MMEJ)可以將兩個斷裂末端連接起來。
因此,在我們預想插入DN**段的位置通過CRISPR/Cas9系統切割生成一個斷裂末端,此刻斷裂點附近如果有DN**段,這個DN**段很可能會被通過末端連接的方式整合進入基因組,這也就是HITI的基本原理。這里提到的DN**段也是一段donor序列,和HDR過程中提供的donor相比,HITI的donor序列只需要包含插入序列,而不需要兩側的同源序列。與HDR對比,HITI由于依賴的是末端連接的修復方式,其效率要高出非常多。且由于不依賴于細胞周期,可以用于終末分化的體細胞的編輯5。
這里簡單介紹一下HITI的donor序列的設計。對于HDR,由于有兩側的同源序列,所以不用擔心插入片段的方向問題。但對于HITI,由于依賴的是末端連接的修復方式,所以如何避免片段被反向連入是需要考慮的問題。作為解決方案,在HITI的donor質粒中引入一個和基因組上靶點相同的序列,但方向和相對于插入序列的方向和基因組上靶點方向相反。
所以該序列也會被sgRNA識別并切割,切割之后如果正向連接進入預期位置,連接位置不會形成新的sgRNA靶點。但如果是反向連接進預期位置,就會和載體上序列重新形成sgRNA靶點,從而被二次識別并切割,從而保證了插入序列的方向性(圖2)。
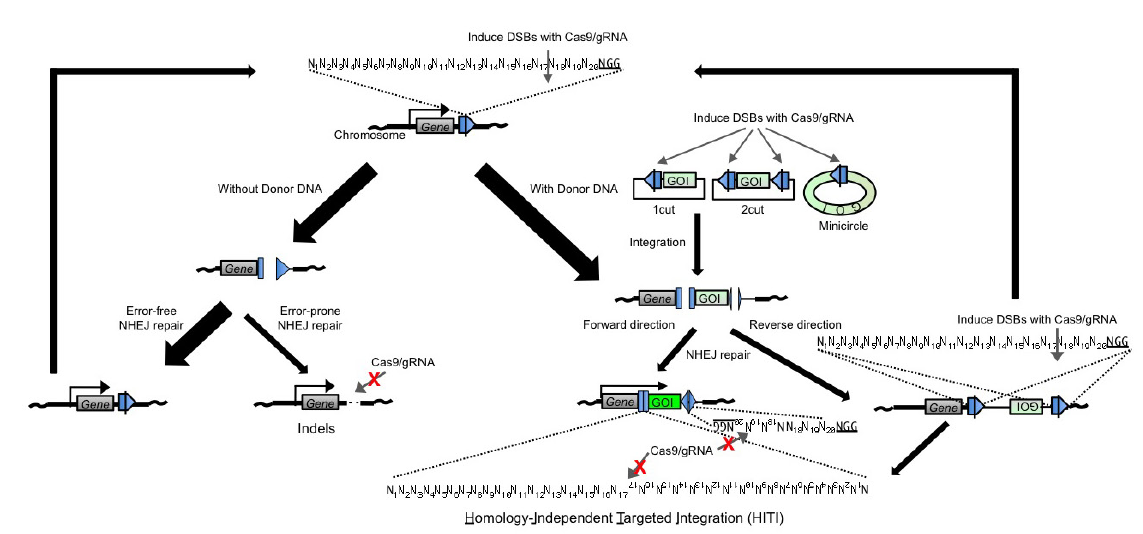
圖 2 通過控制donor質粒上靶點的方向實現插入片段的定向插入
可以看到HITI的修復效率相較于HDR高出非常多(圖3),并且雖然長期認為末端連接的修復方式將引入小片段的插入或者確實(indel),但從實驗結果看,沒有indel的修復結果依然占據了多數。這很可能與NHEJ的修復方式有關系,已有報道證明NHEJ連接兩個沒有修飾的斷裂末端時,會傾向于無indel的修復結果。典型的,如將CtiP敲除,將促進NHEJ,而導致片段敲除中精確連接的比例明顯升高。
因此無論是從修復效率來看,或者是精確修復的絕對比例看,HITI都是更適合體內研究片段插入的技術。
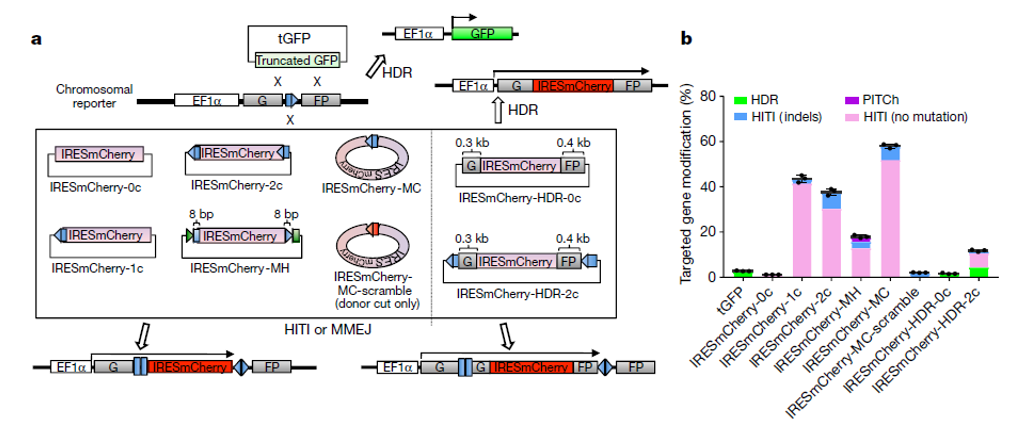
圖 3 HITI修復效率于HDR對比
#微同源介導片段刪除
對于精確的片段插入或者點突變我們都介紹了相對于HDR更高效的方案,那么對于片段的精確刪除是否也有更高效的方案呢?在特定情況下是有的。
我們之前有介紹過,CRISPR/Cas9所介導的DNA雙鏈斷裂的修復結果并不是隨機的(注意!Cas9介導DNA雙鏈斷裂修復結果不是隨機的!)
具有高度可重復性。雖然目前還不確定這種修復結果的有序性具體是如何實現的,但是可以明確是,和細胞本身的DNA雙鏈斷裂修復系統,細胞周期,DNA斷裂點附近序列有高度的相關性。
因此通過大數據分析結合機器學習,在一定程度上可以預測修復后的DNA序列。圖4和圖5介紹了兩個預測修復結果的網站6 7。圖中用于預測的序列具有很強的微同源重復序列,預測結果都顯示近80%比例的基因組在修復后將缺失中間8bp重復序列。
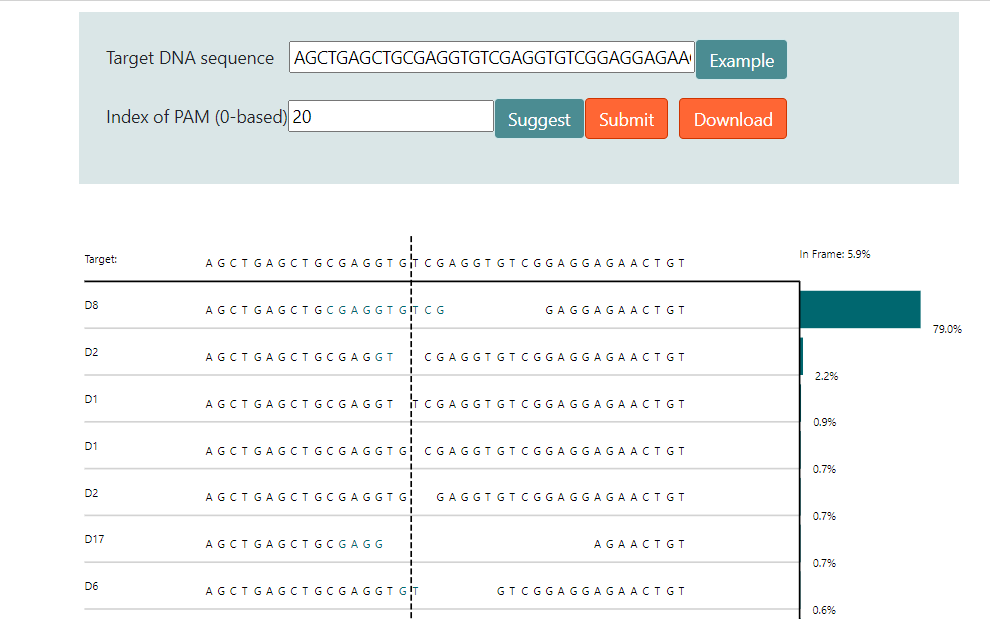
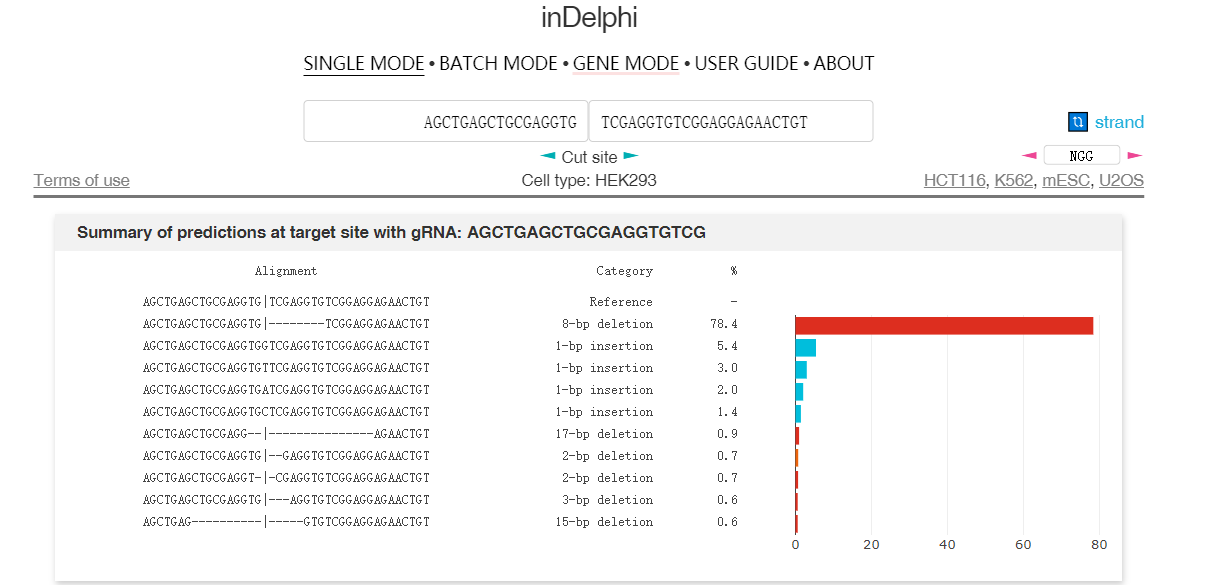
實際上這段序列來源于另外一篇研究MMEJ精確介導重復片段刪除的文章,上面兩個預測結果和文章內實驗結果高度吻合8。TCAP基因中8bp的重復將導致閱讀框移碼從而失活蛋白,最終導致后肢肌肉不良綜合征(LGMD2G)。當在重復序列附近設計sgRNA靶點,通過Cas9切割產生雙鏈斷裂后,細胞將利用MMEJ修復通路刪除重復的序列。
從結果看,在編輯后77%的細胞至少擁有了一個野生型基因(圖6)。這暗示我們,對于擁有某些特定序列的基因組,在雙鏈斷裂后,其修復結果會更傾向于形成單一的修復結果,尤其是部分具有高度微同源序列的DNA。
利用這種特性,我們可以實現特定片段的精確刪除,且具有超高的效率。但是需要注意的一點是,MMEJ和HDR一樣具有細胞周期的依賴性,主要發生在S/G2期,因此如果用該方案去編輯已經退出分裂周期細胞或者終末分化的細胞,將極大地影響效率。
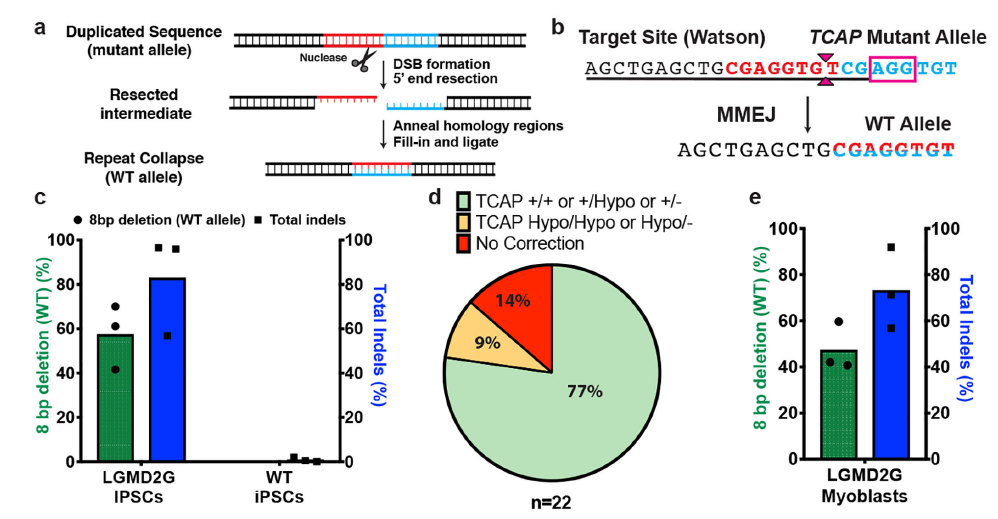
圖 6 MMEJ修復切除重復片段
對于本期介紹到的技術,如果感興趣,歡迎致電吉凱進行咨詢探討。
