
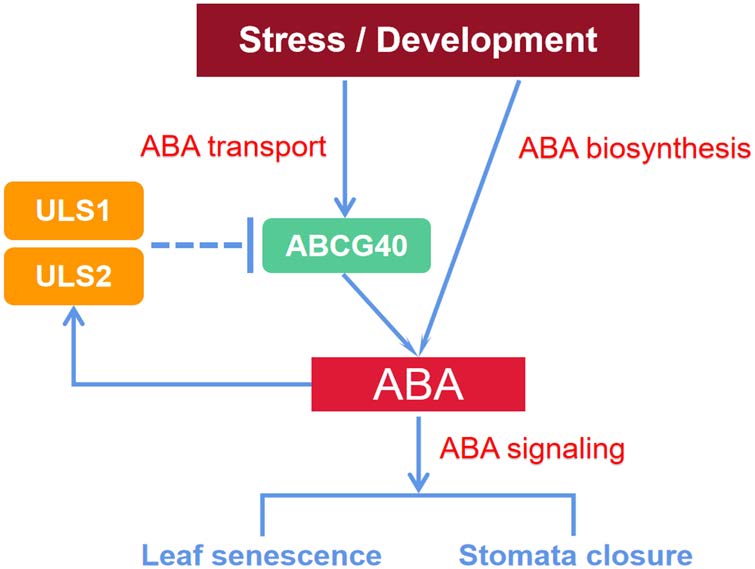
衰老是植物生命周期中不可缺少的過程,而植物激素在葉片衰老過程中起著關鍵作用,其中脫落酸(ABA)、Ethylene、茉莉酸(JA)和水楊酸(SA)是促進葉片衰老的重要物質。其中ABA介導植物的多種生理過程,如氣孔關閉、葉片衰老、種子休眠和滲透調節等。盡管ABA的生物合成和信號轉導以及它們的調控已經得到了廣泛的了解,但ABA是如何通過轉運被調節的還不清楚。
2022年5月,浙江大學丁忠杰教授團隊于Journal of Integrative Plant Biology (IF 9.106)發表了題為“RING‐box proteins regulate leaf senescence and stomatal closure via repression of ABA transporter gene ABCG40”文章,通過轉錄組學及修飾蛋白質組學,研究發現了兩種未被鑒定的RING-box蛋白,衰老泛素連接酶1和2 (ULS1和ULS2),并證明其在ABA信號反饋抑制中作為ABCG40依賴的ABA轉運負調控因子,控制葉片衰老和氣孔關閉,這對ABA轉運調控及其對葉片衰老影響的理解提供了幫助。其中,中科新生命為該研究提供了泛素化修飾蛋白質組學服務。

研究材料
野生型擬南芥及其突變體
技術路線
步驟1:缺乏ULS1和ULS2促進葉片衰老;
步驟2:探究ULS1/2的表達模式;
步驟3:ULS1和ULS2功能的喪失促進ABA誘導的葉片衰老;
步驟4:ULS1/2抑制成熟葉片中ABA的積累;
步驟5:RNA‐seq揭示uls1/uls2中高表達ABA的轉運基因ABCG40;
步驟6:ULS1/2介導的葉片衰老依賴于ABCG40;
步驟7:ULS1/2參與ABCG40依賴的氣孔運動;
步驟8:定量蛋白質組學揭示uls1/uls2中存在差異的泛素化蛋白。
研究結果
1. 缺乏ULS1和ULS2促進葉片衰老
作者首先基于擬南芥現有的基因表達模式,發現兩個屬于RING‐V組的基因在衰老葉片中表達增加,并將其命名為ULS1和ULS2。接著作者通過生成一個雙突變體uls1/uls2,發現其比野生型和單突變型的葉片衰老明顯提前,葉片鮮重和葉綠素含量均降低。此外在此雙突變體中構建原生啟動子驅動的ULSI和ULS2,發現這兩個基因均使uls1/uls2的葉片衰老程度恢復到野生型的水平。這些結果表明,ULS1和ULS2參與抑制擬南芥葉片衰老的過程。
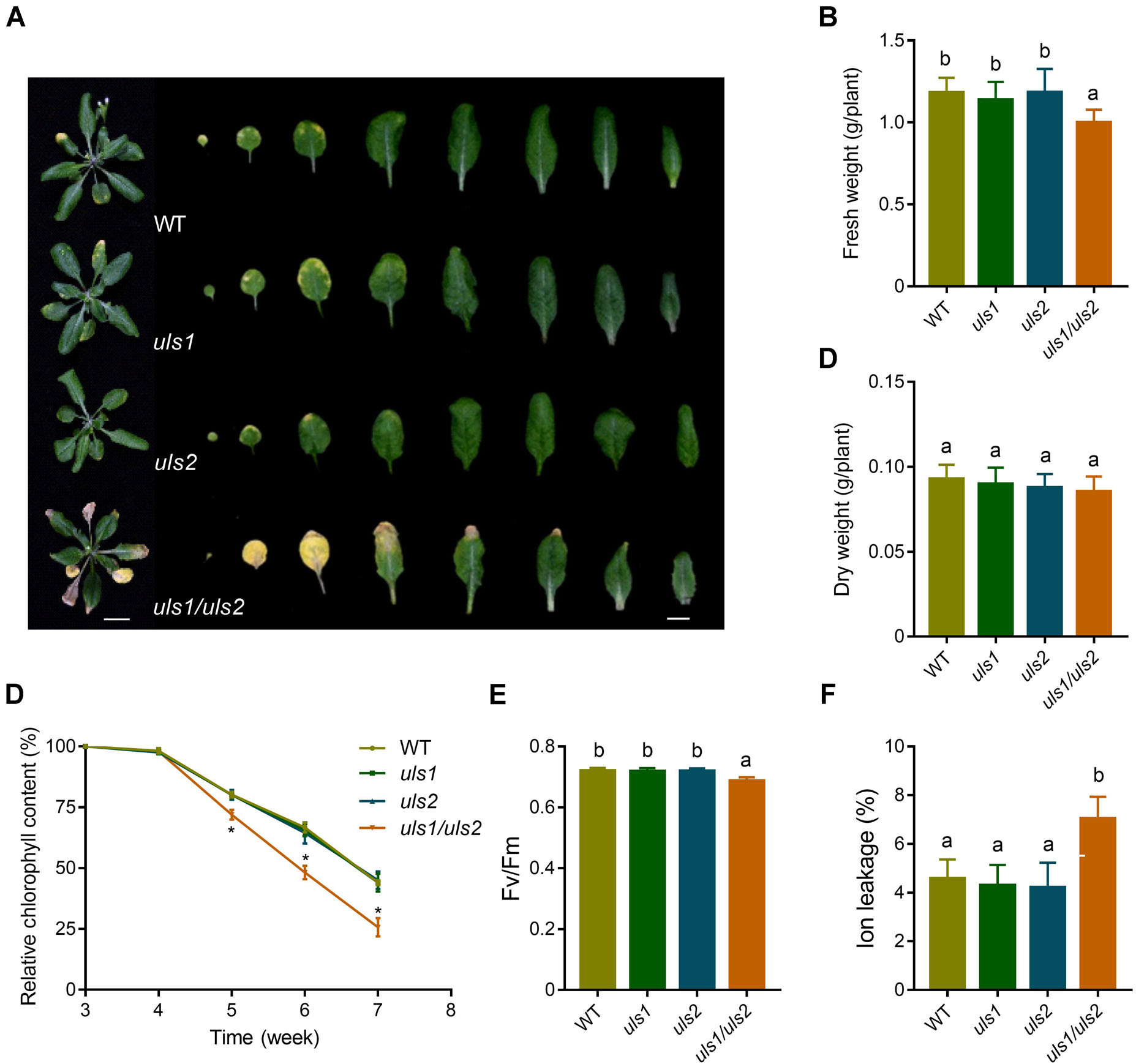
圖1 ULS1和ULS2功能的喪失促進擬南芥葉片衰老
2. ULS1/2的表達模式
作者通過蛋白熒光表達,發現ULS1和ULS2主要在細胞質和內膜系統中表達,并且在葉綠體中顯著富集。為了確定ULS1和ULS2的組織定位,作者構建了由ULS1/2原生啟動子驅動的表達GUS的轉基因株系(PULS1:GUS和PULS2:GUS),通過GUS染色,發現ULS1和ULS2主要表達在莖和根的維管組織中,在老葉中的表達高于幼葉,同時其在保衛細胞、下胚軸及花中也進行表達。此外,作者使用RT‐qPCR檢測發現ULS1和ULS2在未衰老的成熟葉(NL)和老葉(OL)中高度表達,ULS1同樣在花和角果中高度表達,表明其主要在成熟和衰老葉片中表達。
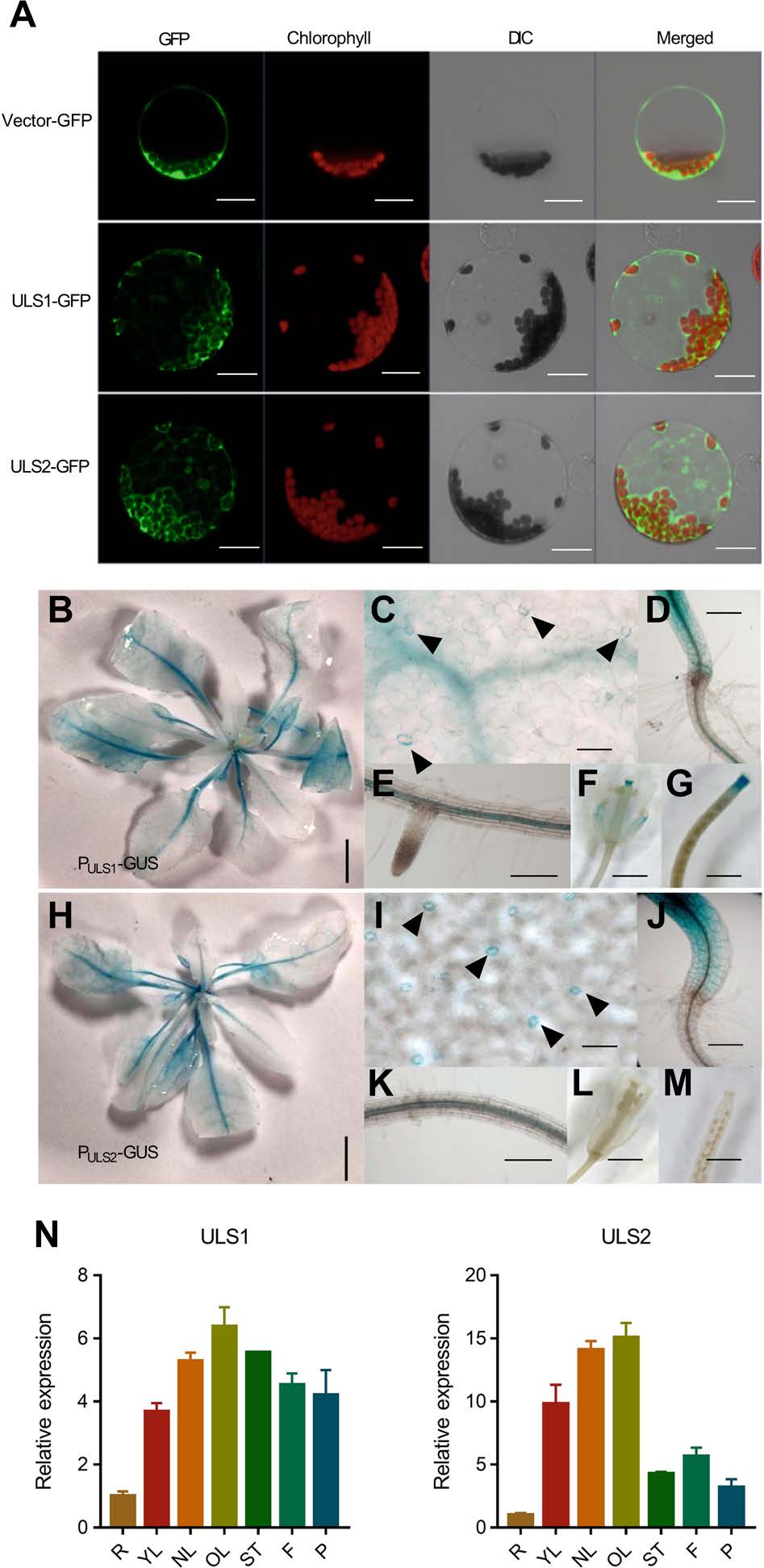
圖2 ULS1和ULS2的表達模式
3. ULS1和ULS2功能的喪失促進ABA誘導的葉片衰老
為了探討ULS1和ULS2是如何參與葉片衰老過程的,作者分別用ABA、SA、MeJA、ACC和黑暗條件處理WT和uls1/uls2突變體的未衰老成熟葉片。結果表明,突變體uls1/uls2葉片衰老加速,葉綠素含量較對照組低。值得關注的是,只有ABA對uls1/ uls2突變體葉片衰老的促進作用明顯大于WT,這表明ULS1和ULS2可能參與了ABA誘導的葉片衰老。為了驗證這一結果,作者測定了WT和uls1/uls2在添加或不添加ABA處理的離體葉片中與衰老相關基因的轉錄水平。結果表明,與WT相比,uls1/ uls2葉片中葉綠素分解代謝基因(NYC1和NYE2)和衰老特異性基因(SAG12和SAG13)的表達顯著增加。此外,ABA能顯著提高uls1/uls2葉片中的基因轉錄水平,其中ULS1和ULS2的mRNA水平明顯受到ABA的誘導。這些數據表明,ULS1和ULS2可能在ABA誘導的葉片衰老過程中起負調控作用。
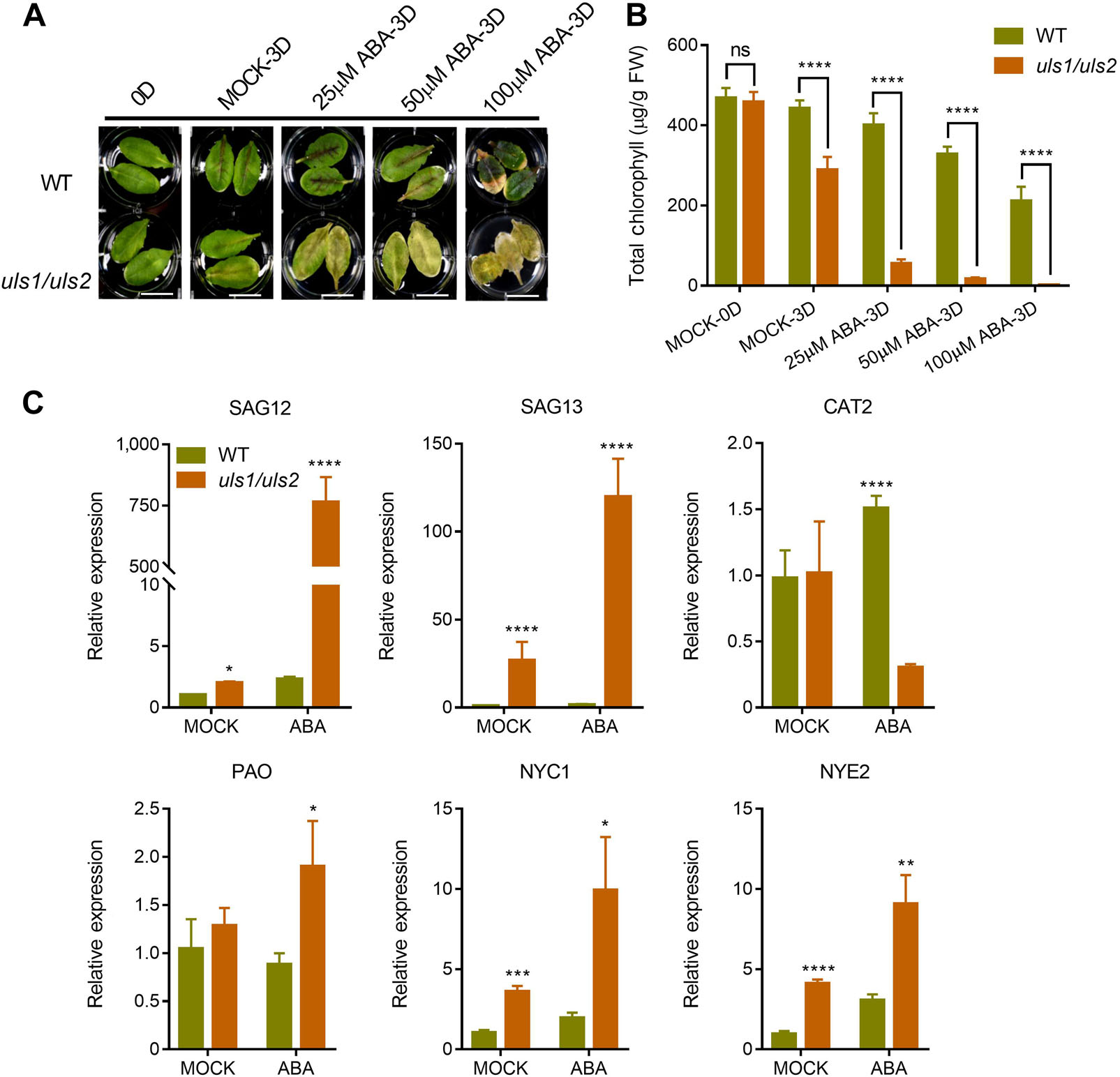
圖3 uls1/uls2對ABA誘導的葉片衰老異常敏感
4. ULS1/2抑制成熟葉片中ABA的積累
為了確定ULS1/2如何影響ABA誘導的葉片衰老,作者將擬南芥葉片分為三個發育階段,即ST1(早熟或幼葉)、ST2(成熟葉)和ST3(老葉),并分別測量了葉片中的ABA含量。結果表明在WT和uls1/uls2植株中,ABA含量都隨著葉片發育而增加。此外,在ST2期,特別是ST3期,uls1/uls2葉片中的ABA積累遠高于WT。這些數據表明,ULS1和ULS2可能通過抑制衰老葉片中ABA的積累來抑制葉片衰老。
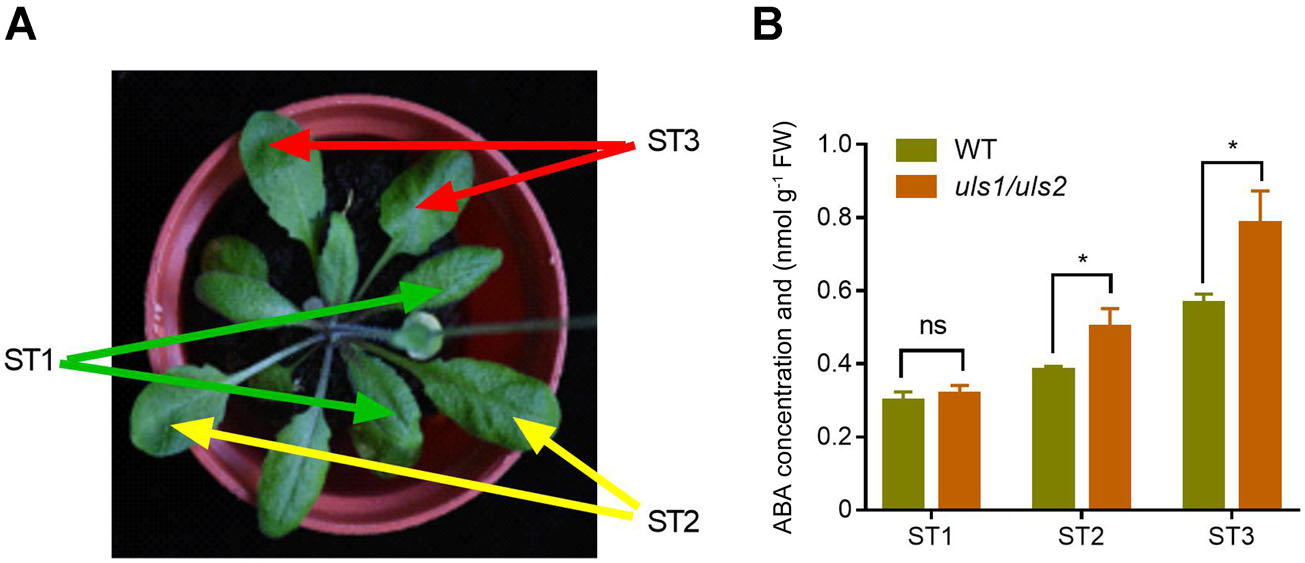
圖4 突變體植株成熟葉中ABA含量增加
5. RNA‐seq揭示uls1/uls2中高表達ABA的轉運基因ABCG40
為了進一步探索ULS1和ULS2調控葉片衰老的分子機制,利用RNA‐seq對WT和uls1/uls2不同階段葉片中差異表達基因進行了鑒定。結果表明,在三個發育階段均有顯著性差異的50個基因中,ABA轉運體基因ABCG40的差異表達最為顯著,并通過RT-qPCR證實了這一結果。此外,在uls1/uls2離體葉片中,ABCG40的表達水平要高得多,且在ABA處理后進一步增強。這些結果表明,ABCG40的高表達可能是uls1/uls2突變體ABA積累和葉片衰老的原因之一。
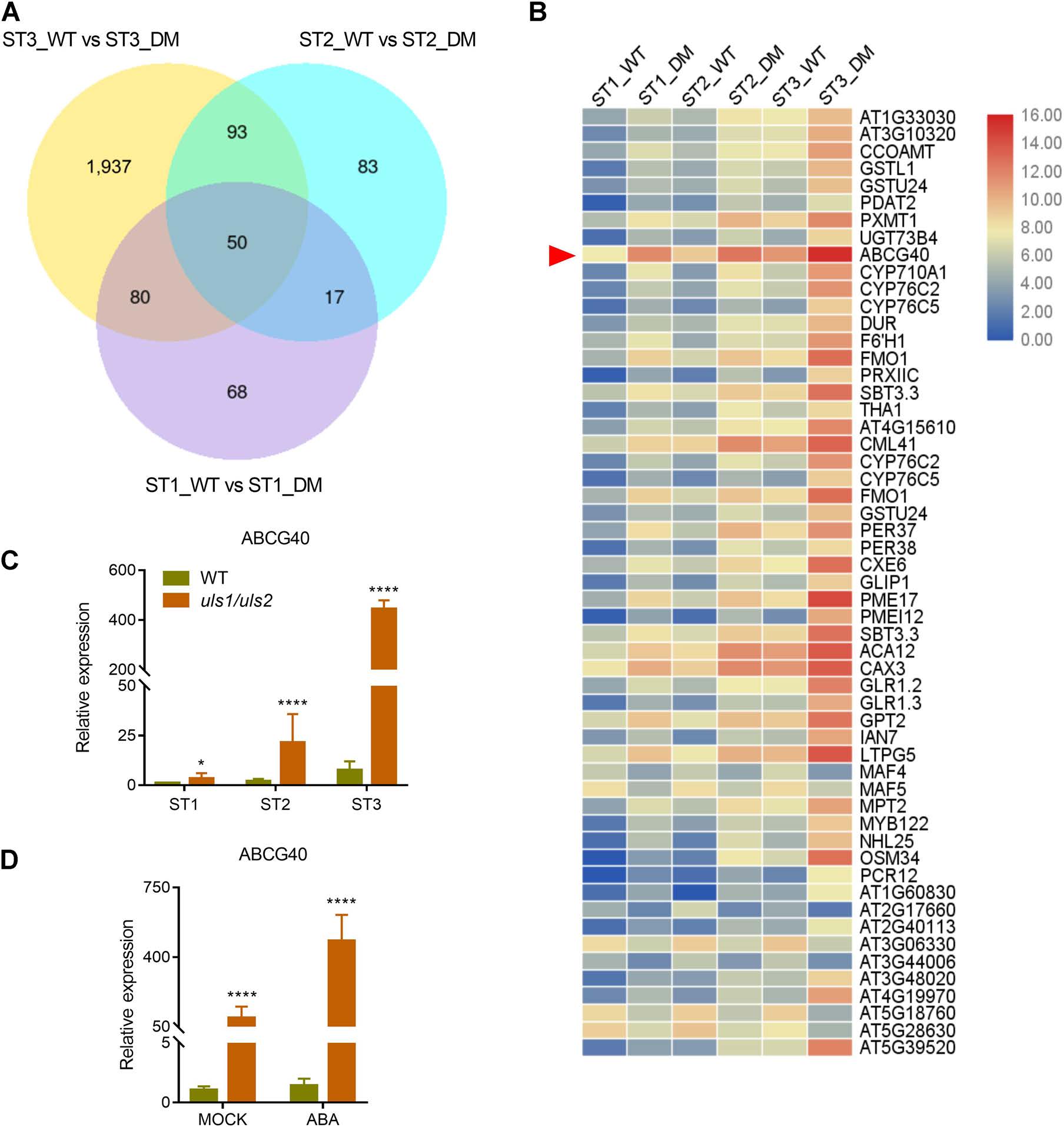
圖5 uls1/uls2葉片在不同發育階段的差異基因表達
6. ULS1/2介導的葉片衰老依賴于ABCG40
緊接著作者開始驗證ULS1和ULS2是否通過ABCG40調節葉片衰老。他們通過構建三重突變體uls1/uls2/abcg40,發現相比較于uls1/uls2,三重突變體在葉片衰老過程中表現出類似WT的表型。此外,與uls1/uls2相比,uls1/uls2/abcg40中衰老相關基因SAG12和NYC1的表達顯著降低。這些結果表明,在缺乏ULS1和ULS2的植物中,ABCG40是導致葉片早期衰老的主要因素。為了進一步證實ABCG40是否有助于增加uls1/uls2中ABA的積累,作者測定了ST3期葉片中的ABA含量,發現uls1/uls2/ ABCG40中ABA的積累明顯低于uls1/uls2。綜上這些數據表明,缺乏ULS1和ULS2會增加ABCG40的表達,進而促進葉片中ABA的積累,導致葉片加速衰老。
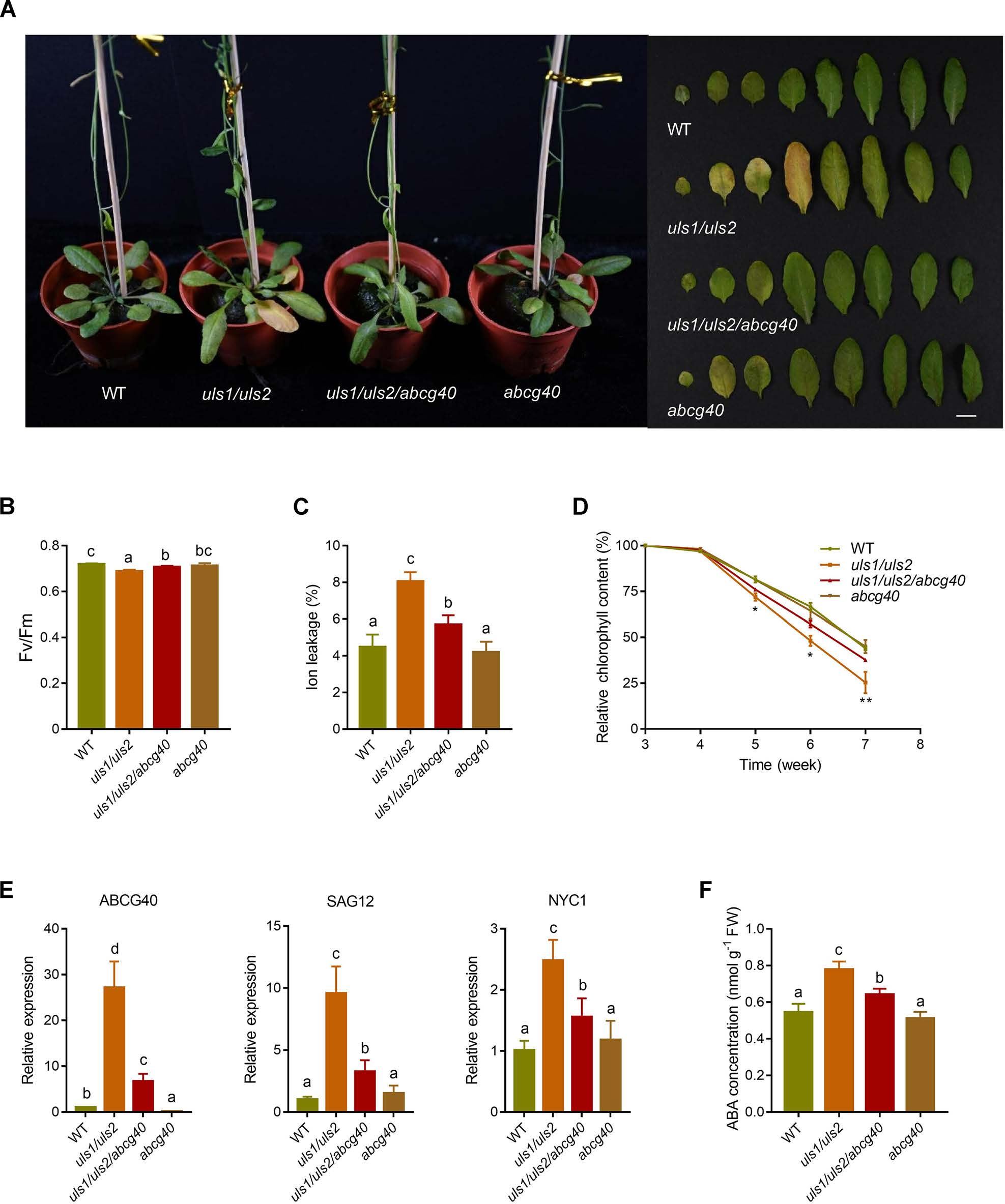
圖6 ULS1和ULS2通過ABCG40參與葉片衰老
7. ULS1/2參與ABCG40依賴的氣孔運動
由于ABA誘導的氣孔關閉需要ABCG40,因此作者接下來探究ULS1和ULS2是否參與氣孔運動的調節。與WT相比,在對照和ABA處理下,uls1/uls2的氣孔孔徑顯著減小,而abcg40突變可以很大程度上抑制uls1/uls2的氣孔關閉,這表明ULS1/2在ABCG40依賴的氣孔運動中起作用。與此一致的是,uls1/uls2的葉片失水率明顯低于WT和uls1/uls2/abcg40突變體。
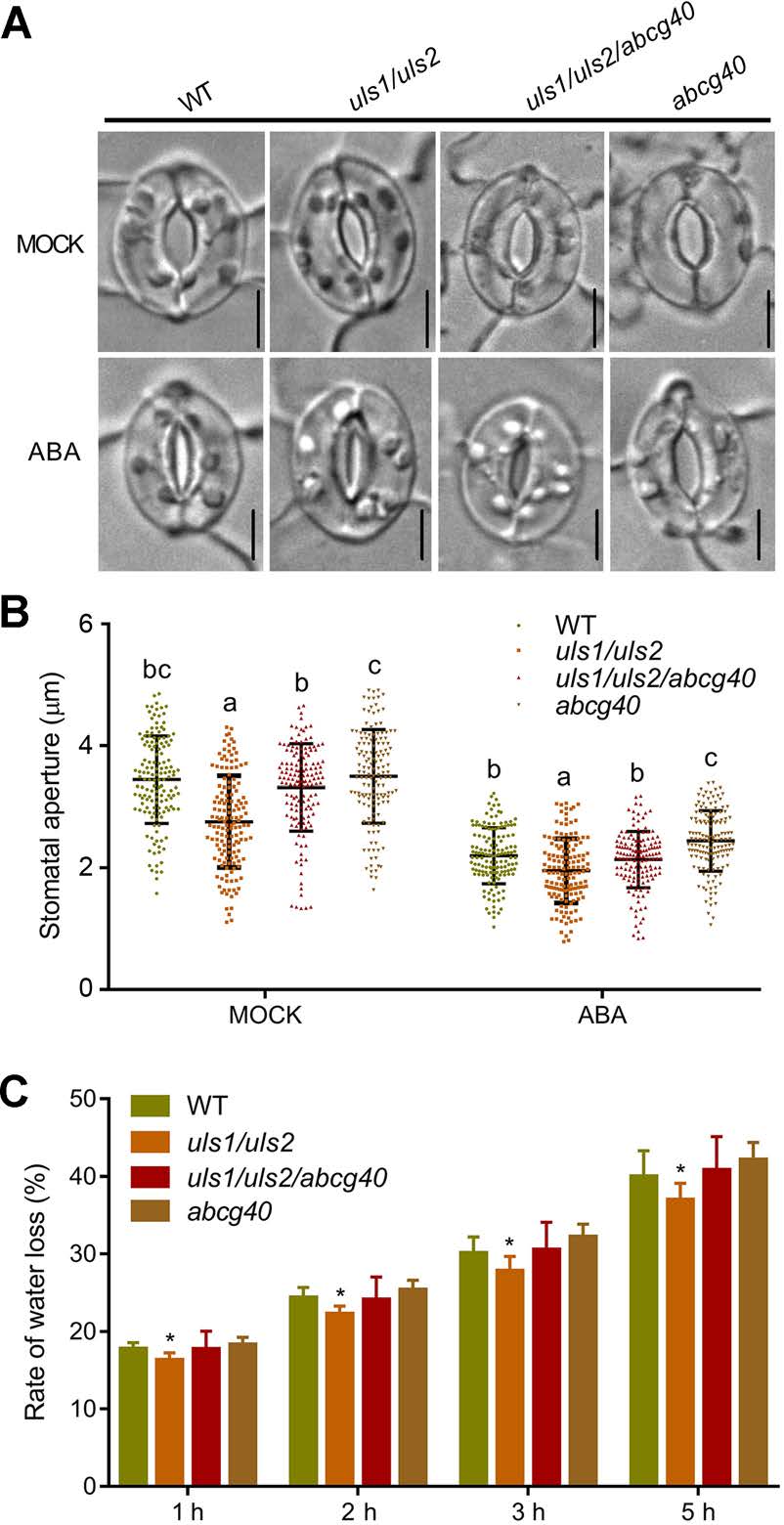
圖7 ULS1和ULS2通過ABCG40調節氣孔孔徑
8. 定量蛋白質組學揭示uls1/uls2中存在差異的泛素化蛋白
接下來文章進一步挖掘ULS1/2如何影響ABCG40的表達。作者從4周齡的WT和uls1/uls2植物中取樣未衰老的成熟葉片,通過Label Free定量蛋白質組學檢測體內泛素化。結果表明共有65個在WT和uls1/uls2之間泛素化差異顯著的蛋白,其中15個蛋白參與ABA響應和衰老過程,支持ULS1/2在ABA誘導的葉片衰老中的作用。此外,ABC轉運蛋白,特別是ABCG40,是uls1/uls2中泛素化水平最高的蛋白之一,表明ABCG40在uls1/uls2植物中豐富,其泛素化受到ULS1/ULS2的間接影響。
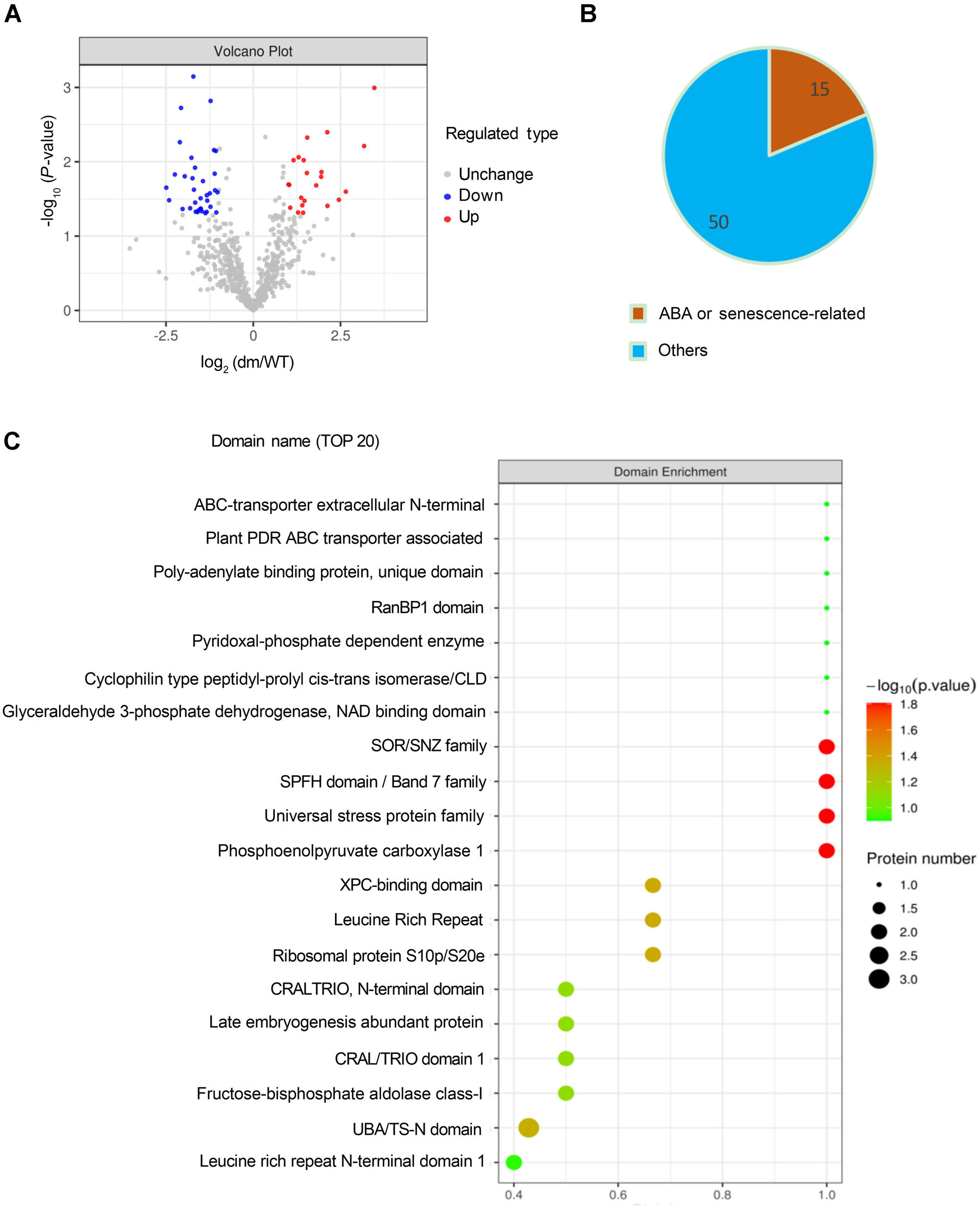
圖8 定量蛋白質組學揭示了WT和uls1/uls2植物中存在差異的泛素化蛋白
小編小結
文章作者研究發現了兩個新的RING家族基因ULS1和ULS2,其通過抑制依賴于ABCG40的ABA運輸來調節葉片衰老和氣孔關閉,為ABA轉運在葉片衰老和氣孔關閉中不可或缺的作用提供了重要證據。