
對食物缺少正常的免疫耐受性與食物過敏(如花生、雞蛋和牛奶)和乳糜瀉是明顯相關。目前尚不清楚保護性免疫是如何被誘導對抗病原體,同時仍保持對食物的免疫耐受的,對這些過程的深入了解將有助于開發針對腸道感染、炎癥和過敏狀況的治療方法。目前免疫耐受的研究主要集中在大腸中的共生細菌上,并涉及抗原呈遞細胞,如CD103+樹突狀細胞(DC)和ILC3s,誘導具有調節功能的T細胞,但對食物免疫耐受的機制研究較少。
2023年7月,中國科學技術大學朱書教授團隊和耶魯大學醫學院 Richard A. Flavell 教授聯合在頂刊Cell(Q1,IF 64.5)上發表了題為“Gasdermin D licenses MHCII induction to maintain food tolerance in small intestine”的文章。該研究發現腸上皮細胞(IECs)積累了一個不易識別的GSDMD的13kD N端片段,該片段在響應膳食抗原時被caspase-3/7裂解。與30kD GSDMD裂解片段不同,IEC積累的GSDMD裂解片段易位到細胞核,誘導CIITA和MHCII分子的轉錄,進而誘導上小腸的Tr1細胞凋亡。用caspase-3/7抑制劑處理的小鼠、抗caspase-3/7裂解的GSDMD突變小鼠、IECs中MHCII缺乏的小鼠和Tr1缺乏的小鼠都表現出食物耐受表型的破壞。所以研究GSDMD的差異裂解可以作為控制小腸免疫與耐受的調節樞紐。中科新生命為該研究提供了蛋白質組學技術服務。

研究材料
小鼠、細胞
技術路線
步驟1:膳食抗原在小腸中誘導非細胞焦亡的13kD GSDMD片段;
步驟2:Caspase-3和caspase-7在IECs中裂解GSDMD;
步驟3:核定位的GSDMD片段協助STAT1誘導IECs中的CIITA和MHCII;
步驟4:核定位的GSDMD片段調節IECs中的MHCII誘導Tr1細胞;
步驟5:在IECs中食物誘導的GSDMD裂解失活會破壞對食物的免疫耐受。
研究結果
1. 膳食抗原在小腸中誘導非細胞焦亡的13kD GSDMD片段
在本研究中,作者用免疫印跡法對腸、胃,淋巴組織和其他器官,包括肺、肝、胰、腦和脊髓進行鑒定,發現全長形式的GSDMD (55 kD),在小腸IEC樣本中意外地觀察到了13kD GSDMD條帶(Figure 1A)。作者想知道GSDMD是否在小腸中發揮某些非細胞焦亡作用。對13kD GSDMD片段進一步研究,利用兩個GSDMD敲除小鼠品系,其中將FLAG-標簽融合到GSDMD的N末端或HA-標簽融合到C末端(Figure 1B和1D)。在這些標記的GSDMD敲除小鼠品系的上小腸中觀察到大小相似的13kD GSDMD條帶(GSDMD-N13)和相應的GSDMD C末端片段(GSDMD-C42)(Figure 1C和1E),說明GSDMD可能裂解為13和42kD片段。接下來,GSDMD-N13片段的信號在IEC中表達明顯(Figure 1F)。這些數據表明,在觸發和功能未知的IEC中,GSDMD可能在小腸上部發生13/42裂解。為了探索觸發小腸上部GSDMD13/42裂解的潛在因素,作者從腸中依次分析了單個環境成分的影響。結果表明抗生物藥(ABX)或抗病毒雞尾酒(AVC)治療、微生物信號或膽汁酸(Figure 1G)和是否有病原體(Figure 1H)均不影響GSDMD-N13和全長GSDMD的分布。考慮到消化道該部位存在豐富的營養物質,推斷食物成分可能誘發了該片段。基于用不同類型的食物喂養小鼠的篩選顯示,與使用不含完整蛋白質的氨基酸食物(AAD)喂養的動物相比,使用正常食物喂養的動物的GSDMD- N13水平顯著更高(Figure 1I),其被特異性地形成為缺乏蛋白質類食物抗原。因此,研究結果表明蛋白性膳食抗原而非共生抗原在上小腸上皮細胞中誘導非細胞焦亡13kD GSDMD片段。
小鼠、細胞
技術路線
步驟1:膳食抗原在小腸中誘導非細胞焦亡的13kD GSDMD片段;
步驟2:Caspase-3和caspase-7在IECs中裂解GSDMD;
步驟3:核定位的GSDMD片段協助STAT1誘導IECs中的CIITA和MHCII;
步驟4:核定位的GSDMD片段調節IECs中的MHCII誘導Tr1細胞;
步驟5:在IECs中食物誘導的GSDMD裂解失活會破壞對食物的免疫耐受。
研究結果
1. 膳食抗原在小腸中誘導非細胞焦亡的13kD GSDMD片段
在本研究中,作者用免疫印跡法對腸、胃,淋巴組織和其他器官,包括肺、肝、胰、腦和脊髓進行鑒定,發現全長形式的GSDMD (55 kD),在小腸IEC樣本中意外地觀察到了13kD GSDMD條帶(Figure 1A)。作者想知道GSDMD是否在小腸中發揮某些非細胞焦亡作用。對13kD GSDMD片段進一步研究,利用兩個GSDMD敲除小鼠品系,其中將FLAG-標簽融合到GSDMD的N末端或HA-標簽融合到C末端(Figure 1B和1D)。在這些標記的GSDMD敲除小鼠品系的上小腸中觀察到大小相似的13kD GSDMD條帶(GSDMD-N13)和相應的GSDMD C末端片段(GSDMD-C42)(Figure 1C和1E),說明GSDMD可能裂解為13和42kD片段。接下來,GSDMD-N13片段的信號在IEC中表達明顯(Figure 1F)。這些數據表明,在觸發和功能未知的IEC中,GSDMD可能在小腸上部發生13/42裂解。為了探索觸發小腸上部GSDMD13/42裂解的潛在因素,作者從腸中依次分析了單個環境成分的影響。結果表明抗生物藥(ABX)或抗病毒雞尾酒(AVC)治療、微生物信號或膽汁酸(Figure 1G)和是否有病原體(Figure 1H)均不影響GSDMD-N13和全長GSDMD的分布。考慮到消化道該部位存在豐富的營養物質,推斷食物成分可能誘發了該片段。基于用不同類型的食物喂養小鼠的篩選顯示,與使用不含完整蛋白質的氨基酸食物(AAD)喂養的動物相比,使用正常食物喂養的動物的GSDMD- N13水平顯著更高(Figure 1I),其被特異性地形成為缺乏蛋白質類食物抗原。因此,研究結果表明蛋白性膳食抗原而非共生抗原在上小腸上皮細胞中誘導非細胞焦亡13kD GSDMD片段。
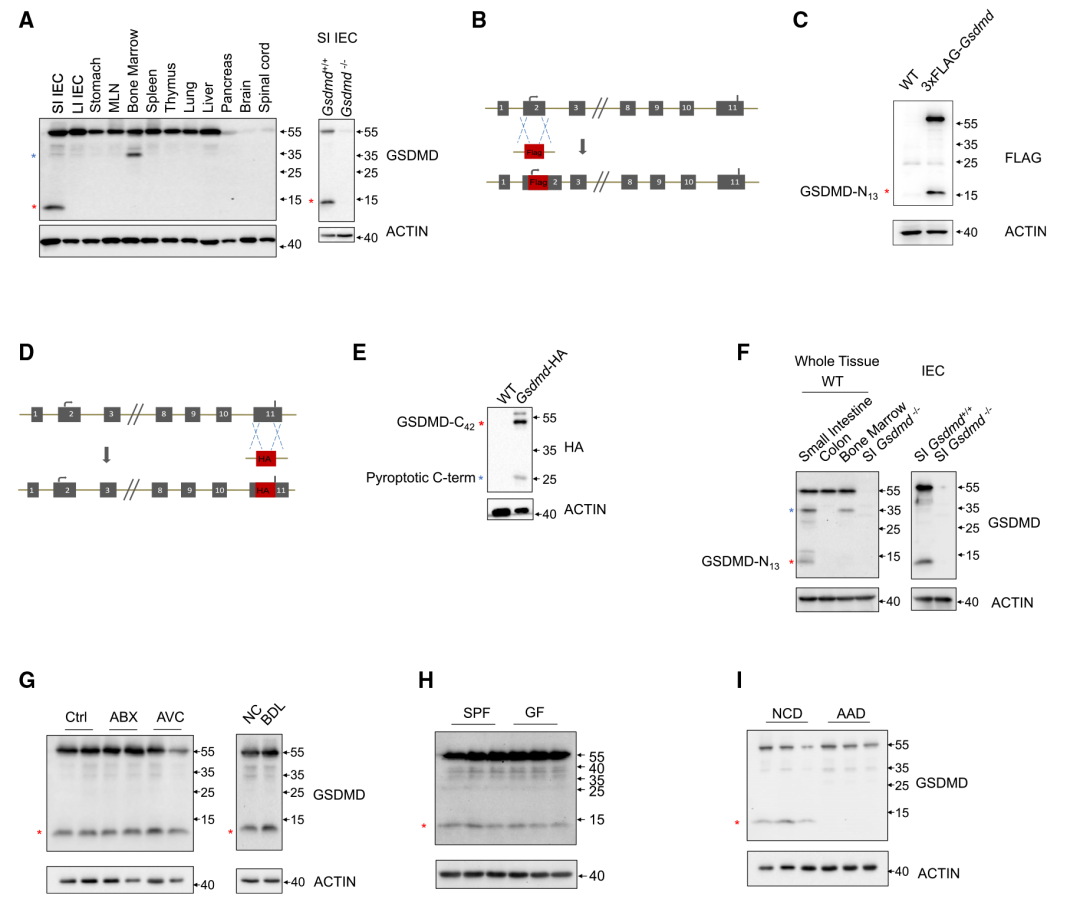
Figure1 膳食抗原在小腸上部誘導非細胞焦亡的13kD GSDMD片段
2.Caspase-3和caspase-7在IECs中裂解GSDMD
作者隨后進行了質譜分析(IP-MS),以確定可能的蛋白酶(一種或多種)裂解GSDMD的13/42,通過對MS結果進行分析,在考馬斯氏染色的SDS-PAGE中,蛋白酶如caspase (CASP)-3、CASP-7和CASP-1被指定為凝膠帶的最佳候選物(Figure 2AB),CASP-1在BM細胞中中度表達(在此處檢測到大量30/25 GSDMD裂解),但在IEC中僅表現出弱表達,CASP-3和CASP-7在IEC中強表達,而在BM細胞中未表達(Figure 2C)。同時還發現CASP-3和CASP-7表達(以及GSDMD-N13片段)發生在小腸上部,包括十二指腸和空腸,一個充滿膳食抗原的區域,不發生在小腸或結腸遠端(Figure 2D)。CASP-7或CASP-3的過表達導致HEK293T細胞中13/42 GSDMD裂解(Figure 2E),用CASP-3/7抑制劑Ac-DEVD-CHO處理共表達GSDMD和CASP-7的HEK-293T細胞,未檢測到GSDMD-N13片段(Figure 2F)。此外,體內實驗顯示,連續4天腹腔內注射Ac-DEVD-CHO可降低來自小腸上部的IEC中CASP-3和CASP-7的表達以及GSDMD的13/42N-末端裂解程度(Figure 2G),證實CASP-3和CASP-7是能夠在體外和體內催化13/42 GSDMD裂解的蛋白酶。
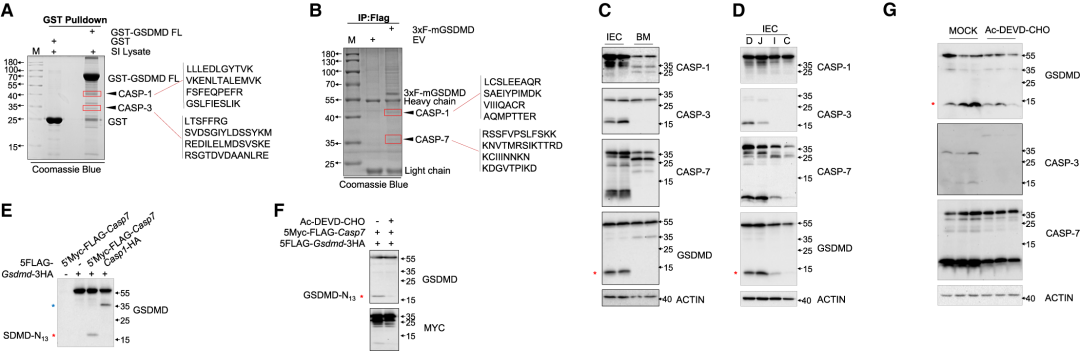
Figure2 Caspase-3和caspase-7在IECs中裂解GSDMD
3. 核定位的GSDMD片段協助STAT1誘導IECs中的CIITA和MHCII
作者進一步研究了IEC中13/42 GSDMD裂解產生的潛在生理變化。首先生成了重組GSDMD的片段,包括響應膳食抗原而裂解的GSDMD- N13片段(1-88 aa),以及相應的GSDMD- C42片段(89-487 aa)。檢測這些重組GSDMD截斷變體的亞細胞定位:GSDMD- C42定位在細胞質中(Figure 3A)。與GSDMD-N30不同,GSDMD-N30定位于引發細胞焦亡的細胞膜,而GSDMD-N13則定位于細胞核(Figure 3A、3B)。此外,在用上述AAD替代正常食物后,檢測到IECs中核定位的GSDMD片段的程度顯著降低,以及CASP-7的激活(Figure 3C),這表明膳食抗原誘導了GSDMD- N13的核易位。
作者通過產生三種小鼠品系(GSDMD完全敲除小鼠(Gsmdmd-/-)、IEC中GSDMD特異性缺陷小鼠(GsdmdΔIEC)和攜帶小腸13/42裂解抗性GSDMD變體的小鼠(GsdmdSICR))探索了哪些途徑可能受到IEC核GSDMD-N13的影響。首先研究團隊分析了來自GSDMD-/-和同窩WT小鼠小腸上部的IEC轉錄組。基因本體(GO)分析表明,在IEC中富集了與抗原加工和呈遞相關的差異調控基因,編碼主要組織相容性復合體II類(MHCII)分子的基因以及被廣泛認為是MHCII分子表達的“主控因子”的Ciita(Figure 3D - 3G)的富集程度尤其高。
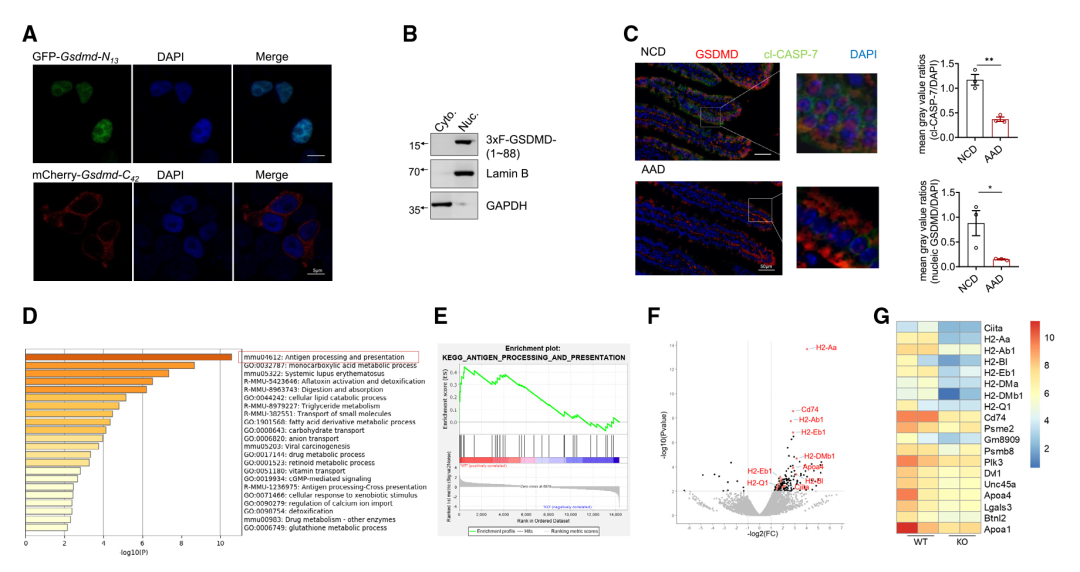
Figure3 核定位的GSDMD片段協助STAT1誘導IECs中的CIITA和MHCII
4. 核定位的GSDMD片段調節IECs中的MHCII誘導Tr1細胞
隨后進行了單細胞RNA測序(scRNA-seq),以分析GsdmdSICR小鼠和相應的仔鼠對照中的基質和免疫細胞群體。上小腸細胞的無監督聚類顯示基于其特定基因表達譜的23個不同聚類(Figure 4A)。Ep-cam(編碼EpCAM)和Ptprc(編碼CD45)的表達分別表明了上皮細胞和免疫細胞,WT IECs顯示了Gsdmd的富集表達(在簇4和簇5中表達最高),還檢測到GsdmdSICR小鼠的幾個細胞簇(如腸細胞和腸干細胞[ISC])中的MHCII分子表達與其WT同窩小鼠相比顯著降低(Figure 4B),并注意到MHCII分子在專門的IEC中幾乎不表達。重要的是為GSDMD-N13與MHCII表達之間的功能關聯提供進一步支持—在穩態下高水平表達MHCII分子的典型抗原致敏細胞類型(如B細胞、巨噬細胞或DC)中,未檢測到突變基因型與WT同窩動物之間的MHCII分子表達發生變化(Figure 4B),從scRNA-seq進一步證實,GSDMD-N13特異性調節穩態IEC中MHCII分子的表達水平,但不調節髓樣細胞中的表達水平。觀察到GSDMD 13/42切割失活導致小腸上部的Tr1細胞(CD4+IL-10+Foxp3-)減少,但未導致Treg細胞(CD4+Foxp3+)減少(Figure 4C),qPCR分析顯示,與WT同窩仔鼠相比,來自GsdmdSICR小鼠小腸上部固有層淋巴細胞(LPL)的分選CD4+ T細胞中Il-10表達顯著降低,與WT同窩仔對照組相比(Figure 4D),由于穩態下小腸中的細胞IL-10蛋白水平無法通過細胞內FACS染色進行評估,作者將IL-10-GFP報告小鼠與GsdmdSICR小鼠或與H2-Ab1ΔIEC小鼠雜交。然后,從所得子代的十二指腸中分離LP,與相應的WT同窩仔鼠相比,在GsdmdSICR小鼠和H2-Ab1-ΔIEC小鼠中CD4+IL-10-GFP+細胞的數量和百分比均顯著降低(Figure 4E和4F)。CD4+Foxp3+和CD4+Foxp3+Rorγt+ 細胞群不受IEC中GSDMD-N13或MHCII分子丟失的影響(Figure 4E和4F)。這些結果表明,IEC中的GSDMD-N13-MHCII軸支持小腸上部的Tr1細胞群。
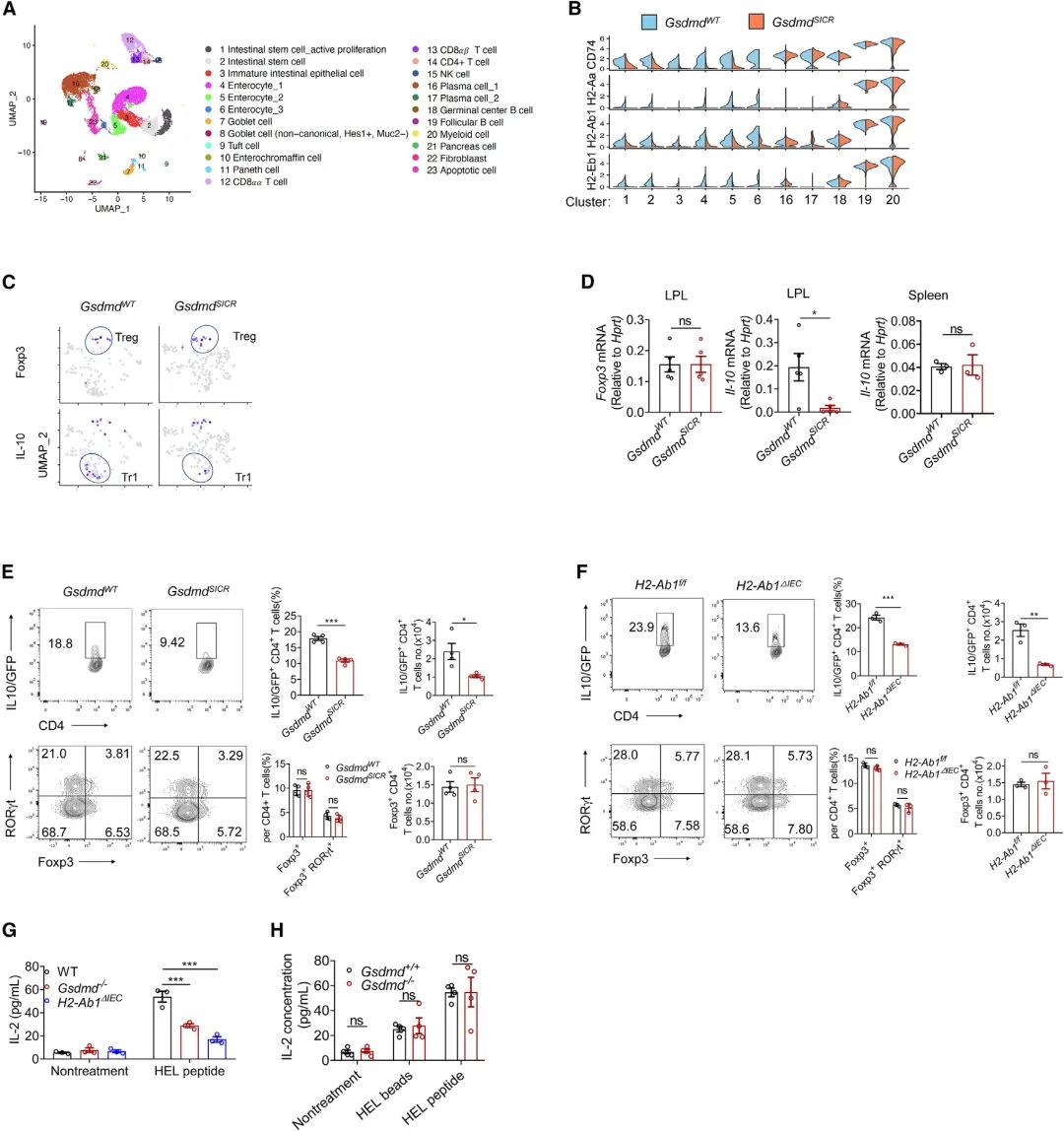
Figure4 核定位的GSDMD片段調節IEC中的MHCII效應以誘導Tr1細胞
5. 在IECs中食物誘導的GSDMD裂解失活會破壞對食物的免疫耐受
進一步探究IEC中的13/42 GSDMD裂解可能參與蛋白質膳食抗原誘導的小腸耐受機制。因此,研究人員用花生提取物誘導了實驗性食物過敏型: (1)GsmdmdΔIEC小鼠對Gsdmdf/f小鼠(即從IEC中去除GSDMD),(2) GsdmdSICR小鼠對GsdmdWT小鼠(阻斷13/42 GSDMD裂解),(3) WT治療小鼠與CASP3/7抑制劑Ac-DEVD-CHO或賦形劑對比對照小鼠(CASP3/7抑制),(4)H2-Ab1ΔIEC小鼠對比H2-Ab1f/f小鼠(去除IEC中MHCII),和(5)因為IL-27是Tr1細胞分化所必需的,因此CD4+ T細胞特異性敲除Wsx1(IL-27R復合物的亞單位)以去除Tr1細胞;因此,與WT對照小鼠相比,研究團隊還在Il27raΔCD4小鼠中誘導食物過敏模型(Figure 5A)。將小鼠暴露于花生蛋白提取物(PN)和霍亂毒素(CT)后,用PN重新攻擊這些小鼠(Figure 5A)中,觀察到所有上述比較組(GsdmdΔIEC小鼠、GsdmdSICR、H2-Ab1ΔIEC、Il27raΔCD4小鼠和Ac-DEVD-CHO處理的小鼠,與它們各自的對照組相比)有以下變化:PN特異性IgE和IgG水平顯著升高,血清mMCP-1水平顯著升高和體溫顯著降低 (Figure 5B-F)。這些變化都是過敏反應的特征,并表明缺乏GSDMD-N13的小鼠、缺乏MHCII的小鼠和缺乏Tr1細胞的小鼠對食物過敏的易感性增加。
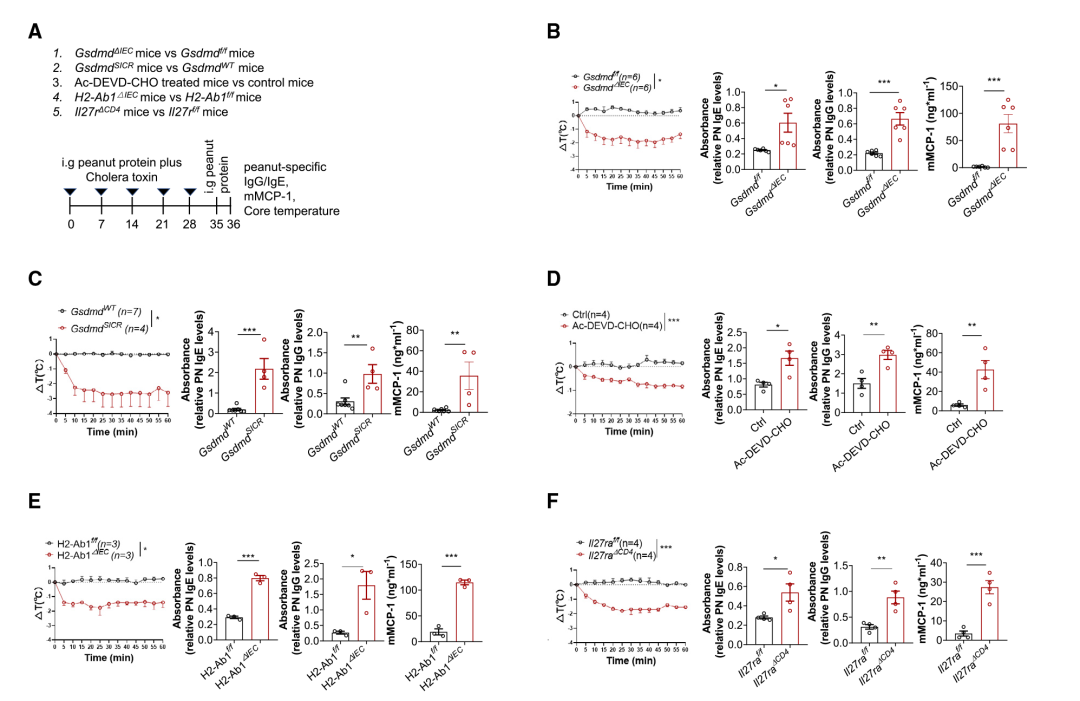
igure5 T GSDMD KO或IECs中抗切割的GSDMD變體會破壞對食物的免疫耐受
文章小結
研究團隊發現IEC在對蛋白質性膳食抗原的反應中積累了非規范的裂解形式的GSDMD,從而通過IEC介導的抗原呈遞支持調節性T輔助亞群Tr1細胞,從而誘導對食物的免疫耐受。這些關于GSDMD在免疫與耐受中的不同作用的深入研究加深了對先天性免疫的理解,并有助于開發治療食物過敏的療法。