自噬相關基因3(ATG3)是一種主要以在LC3脂化過程中發揮作用而聞名的酶。ATG3是否在脂質代謝中發揮作用或參與酒精性脂肪肝病(NAFLD)的發生發展尚不清楚。
來自西班牙的研究團隊在Journal of Hepatology(中科院JCR一區,IF:25.083)發表了題為“Inhibition of ATG3 ameliorates liver steatosis by increasing mitochondrial function”的論文。

研究者通過蛋白質組學檢測了不同小鼠脂肪變性模型中的蛋白變化,并發現了一個與脂質代謝相關的差異變化蛋白——ATG3。ATG3在動物模型和NAFLD患者的肝臟高表達,且可以調控脂質代謝和線粒體活性,是一種與脂肪變性發生發展有關的新蛋白。
研究背景
轉錄因子p63屬于一個由p53、p63和p73組成的家族,是導致肝脂肪變性發展的眾多因素之一。p63編碼多種亞型,可分為2類:具有酸性反轉活化結構域(TA)的亞型和沒有該結構域的亞型(結構域陰性的亞型)。TAp63α亞型在NAFLD動物模型的肝臟和NAFLD肥胖患者的肝臟活檢中均升高。此外,在飲食誘導的肥胖(DIO)小鼠的肝臟中,p63α的下調可減輕肝臟脂肪變性,而TAp63α的激活可通過IKKβ和內質網應激的激活增加肝臟脂肪含量。
本研究,作者通過蛋白質組學檢測了TAp63α基因過表達或敲低的小鼠肝臟中參與脂質代謝的新蛋白。小鼠肝組織蛋白質組學實驗設計:
實驗組1:肝組織特異性過表達TAp53α誘導的脂肪變性小鼠(n=3)
對照組1:正常小鼠 (n=3)
實驗組2:膽堿缺乏高脂食物(CDHFD)誘導的非酒精性脂肪肝小鼠 肝組織特異性敲低p63α(n=3)
對照組2:膽堿缺乏高脂食物(CDHFD)誘導的非酒精性脂肪肝小鼠 肝組織對照(n=3)
研究結果
一、肝臟ATG3受p63的調節,且在飲食誘導的脂肪變性動物模型中高表達
前述兩種實驗設計下的肝臟蛋白質組學分析確定了肝臟脂質積累的新途徑和調節因子。p63正向調控43個蛋白,并顯示了代謝相關蛋白的富集,線粒體為受影響最大的細胞成分。熱圖顯示了一些參與脂質代謝的蛋白,它們受p63過表達的正調控,包括ATG3。考慮到脂吞噬在調節脂肪酸代謝中的相關作用,以及ATG3之前沒有被報道與肝功能相關,作者選擇了這個候選基因進行進一步研究。當肝臟中過表達TAp63α時ATG3 mRNA和蛋白水平上調,而當抑制DIO(飲食誘導肥胖)小鼠肝臟中的TAp63α時,ATG3 mRNA和蛋白水平降低。
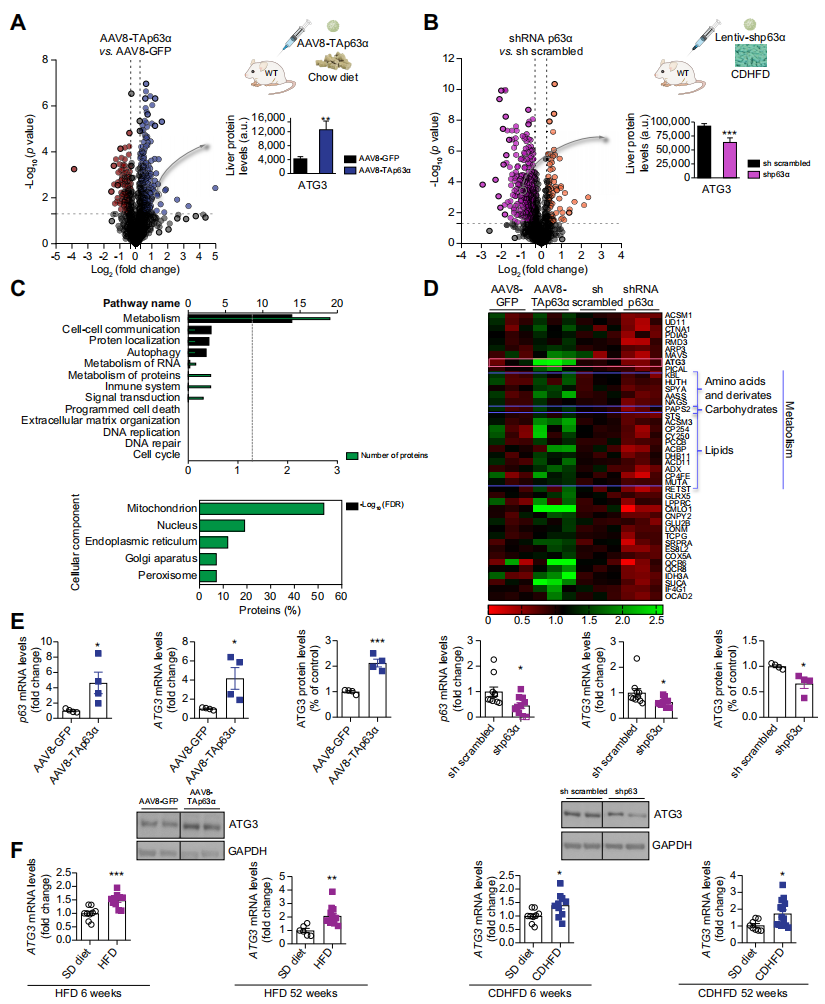
二、NAFLD(非酒精性脂肪肝疾病)患者肝臟中的ATG3升高
在NAFLD患者(25例非酒精性脂肪肝(NAFL)和30例非酒精性脂肪性肝炎(NASH))肝活檢組織中檢測到了比健康肝臟高的ATG3。ATG3與脂肪變性分級、NAFLD活性評分和脂肪肝指數(呈正相關。肥胖(BMI>30)患者與非肥胖(BMI<30)患者相比,ATG3水平升高,并與BMI呈正相關。總之,這些結果表明,在NAFLD患者中存在較高的ATG3水平。
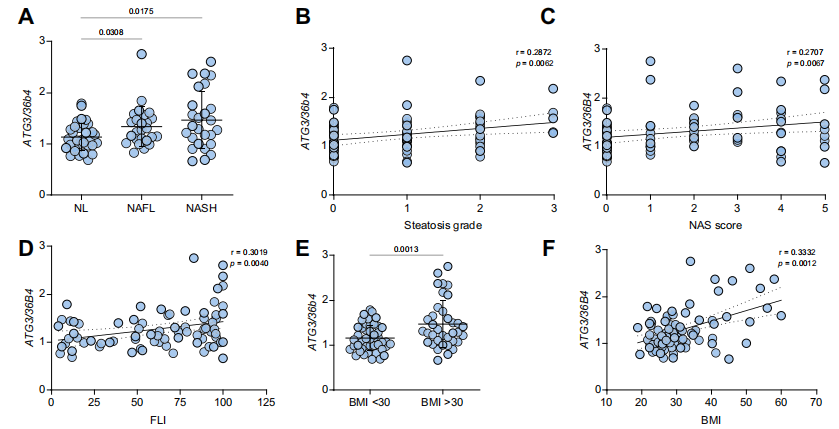
三、ATG3增加了人肝細胞的脂質含量,降低了β-氧化
ATG3 mRNA和脂質含量在經油酸(OA)處理的人肝THLE2細胞增加。然而,ATG3沉默后減少了OA誘導的脂質存儲。如前報道,TAp63α過表達增加了肝細胞中的脂滴,但當ATG3被沉默時,這種作用減弱。在另一種人肝細胞系HepG2中進行的類似實驗也得到了相同的結果。最后,作者從小鼠中分離肝細胞并用OA處理,在轉染siATG3和磷酸化的ATG3后,分別檢測到了脂肪酸β-氧化的增加和減少。
四、ATG3調節肝細胞線粒體活性
在HepG2細胞中過表達ATG3降低了耗氧率。更具體地說,ATG3過表達后,基礎呼吸、ATP相關呼吸、質子泄漏、最大呼吸和24小時非線粒體耗氧量均降低。這些結果與線粒體復合物II(II-SDHB)、III和V的蛋白水平降低相一致。脂肪酸合酶(FAS)、乙酰輔酶A羧化酶或肉堿棕櫚酰基轉移酶1a(CPT1a)的蛋白水平均無變化。然而,ATG3的誘導降低了過氧化物酶體增殖物激活受體伽馬共激活物1α(PGC1a)和sirtuin1(SIRT1),這兩種是促進線粒體功能和脂肪酸氧化的蛋白。HepG2細胞中的ATG3抑制得到了相反的結果。以上這些結果也在THLE2細胞中得到了證實。
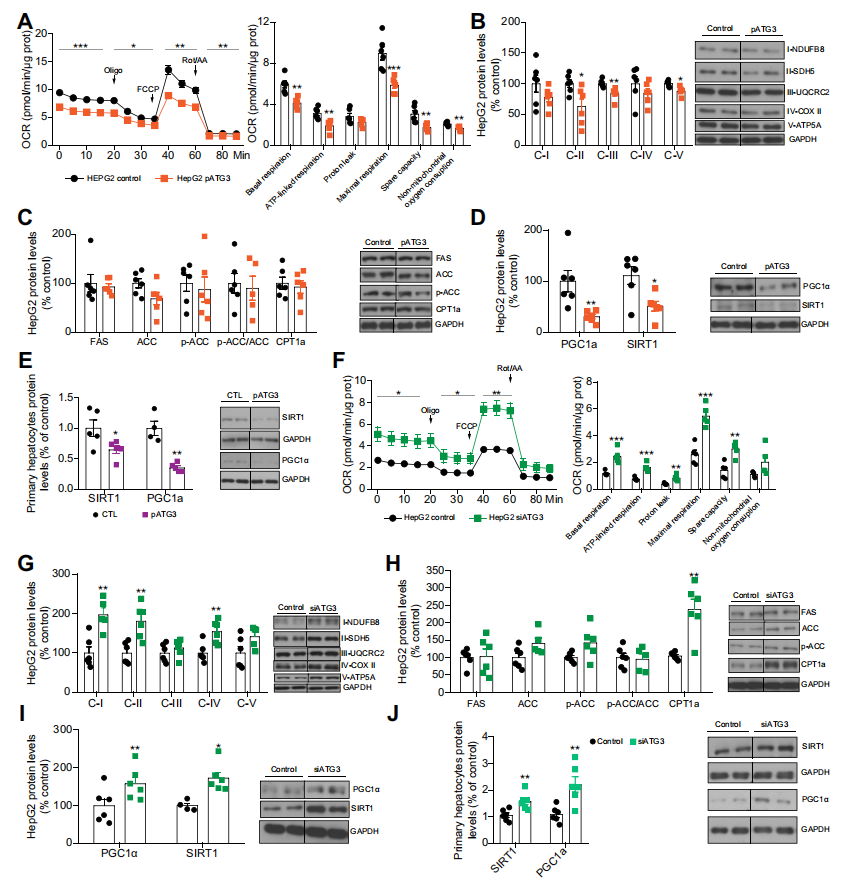
五、抑制肝臟ATG3可改善TAp63α誘導的脂肪變性
作者接下來評估了抑制ATG3對TAp63α誘導的小鼠脂肪變性的影響。TAp63α過表達后,循環天冬氨酸轉氨酶(AST)和丙氨酸轉氨酶(ALT)水平升高。抑制ATG3可降低AST和ALT水平。TAp63α的過表達導致肝臟脂肪變性,這種作用被ATG3敲低所減弱。在機制上,TAp63α過表達的小鼠脂肪酸合成酶(FAS)水平升高,CPT1a、PGC1a和SIRT1水平降低。抑制ATG3可使CPT1a、PGC1a和SIRT1的水平恢復到對照組的水平。肝臟過表達TAp63α降低了所有線粒體復合物的水平,而抑制ATG3則使肝臟復合物I和II的水平恢復正常。
六、ATG3需要SIRT1來調節脂質代謝和線粒體活性
接下來,作者評估了SIRT1作為ATG3的潛在調節因子的功能相關性。在HepG2細胞中沉默ATG3可以改善OA誘導的脂質負荷。然而,當SIRT1也被沉默時,干擾ATG3未能降低OA誘導的脂質含量。這些結果也在共轉染siATG3和siSIRT1的THLE2細胞中得到了證實。此外,在SIRT1沉默后,siATG3在THLE2細胞中誘導的較高的耗氧速率也消失了。SIRT1被白藜蘆醇藥理激活后,ATG3誘導的脂質含量減少。在分離的肝細胞中,ATG3也降低了耗氧率,而白藜蘆醇則減弱了這種作用。與這些結果相一致,沉默ATG3增加了SIRT1的活性,降低了PGC1a的乙酰化水平,而ATG3過表達則降低了分離的肝細胞中SIRT1的活性。
體內實驗中,作者對喂食CDHFD的小鼠先后沉默ATG3和SIRT1。在ATG3基因敲低后,AST和ALT均降低,而當ATG3和SIRT1均被沉默時,這種降低被消除。當SIRT1被抑制時,ATG3下調小鼠脂質含量和肝臟甘油三酯的功能被減弱。肝臟ATG3的敲低導致了CPT1a蛋白水平的增加以及復合物II活性的提高,但這些作用被共沉默ATG3和SIRT1所阻斷。因此,ATG3需要SIRT1來調節脂質代謝和線粒體活性。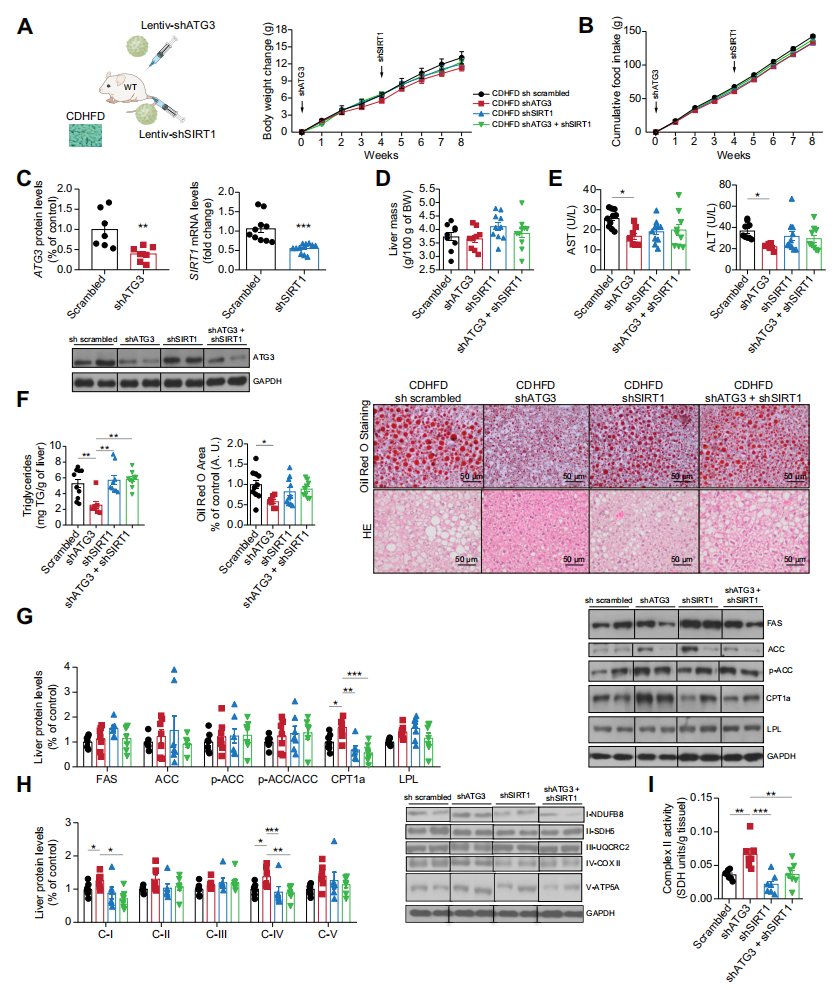
七、ATG3通過JNK1調控SIRT1蛋白水平
調控SIRT1蛋白水平的關鍵因子是c-Jun N端激酶1(JNK1),其激活誘導SIRT1蛋白降解,而JNK2誘導SIRT1蛋白穩定性。作者檢測了用CDHFD喂養的小鼠肝臟中的JNK1和JNK2,在肝臟中ATG3被敲除,檢測到JNK1水平降低,而JNK2水平沒有降低。細胞數據與體內實驗數據一致。最后,用JNK1抑制劑SP600125處理分離的肝細胞,可減弱ATG3誘導的作用,包括增加脂質含量、降低耗氧率和SIRT1活性。
八、ATG3需要CPT1a來調節脂質代謝和線粒體活性
CPT1a floxed小鼠注射AAV8-GFP(對照)或AAV8-Cre;第4周,實驗組注射干擾shRNA(對照)或shRNA-ATG3,第8周犧牲掉小鼠。經AAV8-Cre或shRNA-ATG3處理后,肝臟中的CPT1a和ATG3水平分別降低,而肝臟質量不受影響。但在ATG3基因敲低后,AST和ALT水平降低。當ATG3和CPT1a均被沉默時,它們恢復到基線水平。當CPT1a敲低后,原本ATG3下調導致的小鼠脂質和肝臟甘油三酯降低以及復合物II和IV水平的升高被削弱。這些結果表明,ATG3需要CPT1a來調節脂質代謝和線粒體活性。
九、ATG3以不依賴自噬的方式調節肝脂質含量
作者進一步探索ATG3是否通過自噬依賴的方式發揮作用。在自噬誘導后,微管相關蛋白1輕鏈3α(MAP1LC3A或LC3)被脂化,LC3-磷脂偶聯物(LC3-II)被招募到自噬體膜,與溶酶體融合形成自溶酶體,降解自噬體內成分和LC3-II。與預期的一樣,在HepG2細胞沉默ATG3則降低了LC3-II水平。緊接著,在存在或不存在溶酶體介導的蛋白質水解抑制劑氯喹(CQ)的情況下,作者通過分析LC3-II周轉來監測自噬通量。CQ給藥誘導了預期的LC3-II積累。用OA處理和轉染siATG3的肝細胞儲存了較少的脂質,而CQ處理增加了脂質含量。然而,在過表達ATG3的細胞中,CQ抑制自噬并不影響ATG3誘導的THLE2或HepG2細胞中的脂滴。因此,這些結果表明,ATG3誘導的脂質積累不依賴于自噬。這在體內實驗中得到了證實,因為在兩個脂肪變性動物模型(由TAp63α過表達或CDHFD誘導)中,ATG3敲低減少了LC3-II的積累,但不影響成熟的自噬標記物如ATG5、ATG7和p62的蛋白和mRNA水平。總之,這些數據表明ATG3對脂質含量的影響獨立于自噬作用。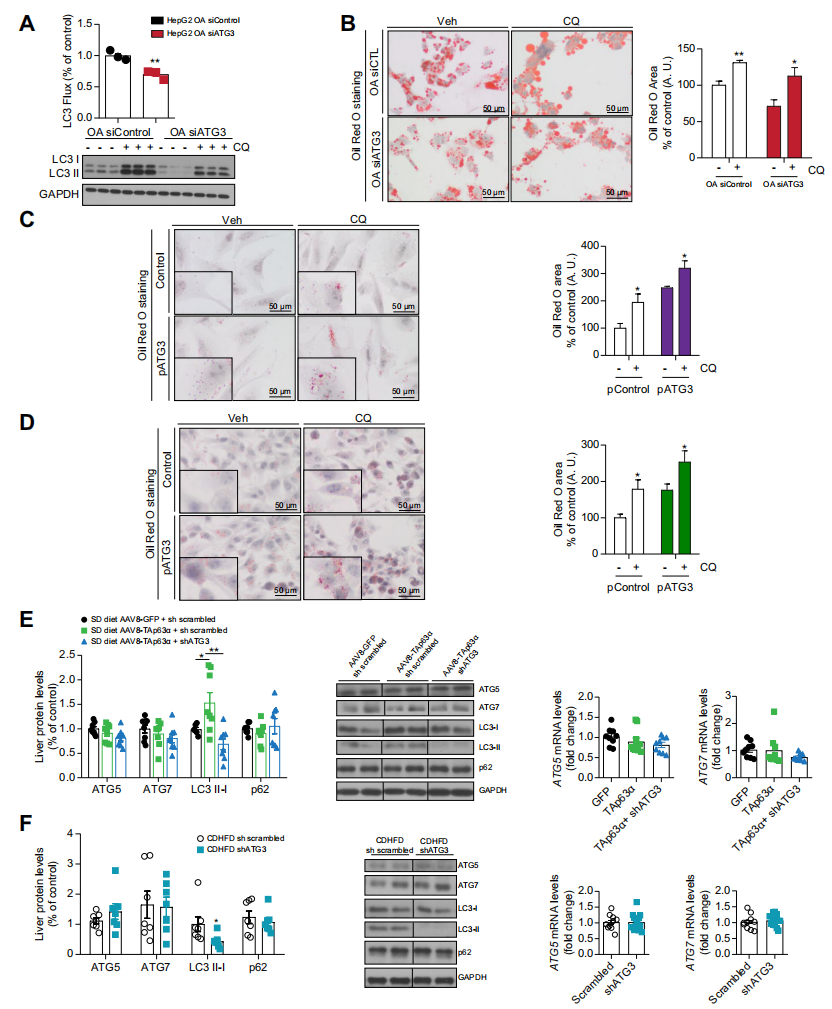
研究總結
作者使用蛋白質組學檢測了TAp63α過表達誘導的小鼠脂肪變性模型的蛋白變化以及CDHFD誘導的小鼠脂肪變性模型在TAp63敲低后的蛋白表達變化。研究發現自噬相關基因3(ATG3)通過TAp63α的激活而上調,而在TAp63α的抑制后則被下調。ATG3在一些NAFLD動物模型和NAFLD患者的肝臟中升高。ATG3基因過表達增加了肝細胞的脂質負荷,而其抑制則減輕了TAp63α和飲食誘導的脂肪變性。ATG3通過調節SIRT1和線粒體功能發揮脂質代謝作用。總體來說,作者的研究確定了ATG3是一個與脂肪變性發展有關的新基因。
吉凱基因憑借多年在靶標篩選及驗證服務領域的技術積累,建立的標準化 、工程化 、系統化的GRP平臺,為中國研究型醫生提供科研服務,加快科研成果轉化。其中,多組學平臺包含蛋白質組學平臺和單細胞測序平臺:
·蛋白質組學平臺擁有多臺timsTOF Pro、Exploris 480高精度質譜儀,專業的Spectronaut Plusar、Mascot等分析軟件,提供專業的4D、DIA、TMT、PRM、磷酸化修飾組和Olink蛋白質組等檢測服務,強大的機器學習算法、IPA分析、蛋白基因組分析服務,系統的生物標志物、分子分型、藥物靶點、基因功能研究等解決方案,真正讓廣大研究型醫生的科研工作更省心、更省力、更高效;
·單細胞測序擁有10x和BD兩個平臺,提供單細胞RNA-seq、單細胞核測序、單細胞混樣RNA-seq、單細胞(RNA+ATAC)、空間轉錄組測序等服務。
接下來,作者評估了SIRT1作為ATG3的潛在調節因子的功能相關性。在HepG2細胞中沉默ATG3可以改善OA誘導的脂質負荷。然而,當SIRT1也被沉默時,干擾ATG3未能降低OA誘導的脂質含量。這些結果也在共轉染siATG3和siSIRT1的THLE2細胞中得到了證實。此外,在SIRT1沉默后,siATG3在THLE2細胞中誘導的較高的耗氧速率也消失了。SIRT1被白藜蘆醇藥理激活后,ATG3誘導的脂質含量減少。在分離的肝細胞中,ATG3也降低了耗氧率,而白藜蘆醇則減弱了這種作用。與這些結果相一致,沉默ATG3增加了SIRT1的活性,降低了PGC1a的乙酰化水平,而ATG3過表達則降低了分離的肝細胞中SIRT1的活性。
體內實驗中,作者對喂食CDHFD的小鼠先后沉默ATG3和SIRT1。在ATG3基因敲低后,AST和ALT均降低,而當ATG3和SIRT1均被沉默時,這種降低被消除。當SIRT1被抑制時,ATG3下調小鼠脂質含量和肝臟甘油三酯的功能被減弱。肝臟ATG3的敲低導致了CPT1a蛋白水平的增加以及復合物II活性的提高,但這些作用被共沉默ATG3和SIRT1所阻斷。因此,ATG3需要SIRT1來調節脂質代謝和線粒體活性。
七、ATG3通過JNK1調控SIRT1蛋白水平
調控SIRT1蛋白水平的關鍵因子是c-Jun N端激酶1(JNK1),其激活誘導SIRT1蛋白降解,而JNK2誘導SIRT1蛋白穩定性。作者檢測了用CDHFD喂養的小鼠肝臟中的JNK1和JNK2,在肝臟中ATG3被敲除,檢測到JNK1水平降低,而JNK2水平沒有降低。細胞數據與體內實驗數據一致。最后,用JNK1抑制劑SP600125處理分離的肝細胞,可減弱ATG3誘導的作用,包括增加脂質含量、降低耗氧率和SIRT1活性。
八、ATG3需要CPT1a來調節脂質代謝和線粒體活性
CPT1a floxed小鼠注射AAV8-GFP(對照)或AAV8-Cre;第4周,實驗組注射干擾shRNA(對照)或shRNA-ATG3,第8周犧牲掉小鼠。經AAV8-Cre或shRNA-ATG3處理后,肝臟中的CPT1a和ATG3水平分別降低,而肝臟質量不受影響。但在ATG3基因敲低后,AST和ALT水平降低。當ATG3和CPT1a均被沉默時,它們恢復到基線水平。當CPT1a敲低后,原本ATG3下調導致的小鼠脂質和肝臟甘油三酯降低以及復合物II和IV水平的升高被削弱。這些結果表明,ATG3需要CPT1a來調節脂質代謝和線粒體活性。
九、ATG3以不依賴自噬的方式調節肝脂質含量
作者進一步探索ATG3是否通過自噬依賴的方式發揮作用。在自噬誘導后,微管相關蛋白1輕鏈3α(MAP1LC3A或LC3)被脂化,LC3-磷脂偶聯物(LC3-II)被招募到自噬體膜,與溶酶體融合形成自溶酶體,降解自噬體內成分和LC3-II。與預期的一樣,在HepG2細胞沉默ATG3則降低了LC3-II水平。緊接著,在存在或不存在溶酶體介導的蛋白質水解抑制劑氯喹(CQ)的情況下,作者通過分析LC3-II周轉來監測自噬通量。CQ給藥誘導了預期的LC3-II積累。用OA處理和轉染siATG3的肝細胞儲存了較少的脂質,而CQ處理增加了脂質含量。然而,在過表達ATG3的細胞中,CQ抑制自噬并不影響ATG3誘導的THLE2或HepG2細胞中的脂滴。因此,這些結果表明,ATG3誘導的脂質積累不依賴于自噬。這在體內實驗中得到了證實,因為在兩個脂肪變性動物模型(由TAp63α過表達或CDHFD誘導)中,ATG3敲低減少了LC3-II的積累,但不影響成熟的自噬標記物如ATG5、ATG7和p62的蛋白和mRNA水平。總之,這些數據表明ATG3對脂質含量的影響獨立于自噬作用。
研究總結
作者使用蛋白質組學檢測了TAp63α過表達誘導的小鼠脂肪變性模型的蛋白變化以及CDHFD誘導的小鼠脂肪變性模型在TAp63敲低后的蛋白表達變化。研究發現自噬相關基因3(ATG3)通過TAp63α的激活而上調,而在TAp63α的抑制后則被下調。ATG3在一些NAFLD動物模型和NAFLD患者的肝臟中升高。ATG3基因過表達增加了肝細胞的脂質負荷,而其抑制則減輕了TAp63α和飲食誘導的脂肪變性。ATG3通過調節SIRT1和線粒體功能發揮脂質代謝作用。總體來說,作者的研究確定了ATG3是一個與脂肪變性發展有關的新基因。
吉凱基因憑借多年在靶標篩選及驗證服務領域的技術積累,建立的標準化 、工程化 、系統化的GRP平臺,為中國研究型醫生提供科研服務,加快科研成果轉化。其中,多組學平臺包含蛋白質組學平臺和單細胞測序平臺:
·蛋白質組學平臺擁有多臺timsTOF Pro、Exploris 480高精度質譜儀,專業的Spectronaut Plusar、Mascot等分析軟件,提供專業的4D、DIA、TMT、PRM、磷酸化修飾組和Olink蛋白質組等檢測服務,強大的機器學習算法、IPA分析、蛋白基因組分析服務,系統的生物標志物、分子分型、藥物靶點、基因功能研究等解決方案,真正讓廣大研究型醫生的科研工作更省心、更省力、更高效;
·單細胞測序擁有10x和BD兩個平臺,提供單細胞RNA-seq、單細胞核測序、單細胞混樣RNA-seq、單細胞(RNA+ATAC)、空間轉錄組測序等服務。