自噬-溶酶體途徑是真核生物細(xì)胞內(nèi)用來(lái)抵抗病原菌,清除變性蛋白和異常細(xì)胞器的主要機(jī)制之一。自噬-溶酶體途徑的異常和包括帕金森癥、阿茲海默癥在內(nèi)的多種神經(jīng)退行性疾病以及癌癥密切相關(guān)。TFEB 是非常重要的轉(zhuǎn)錄因子,調(diào)控自噬相關(guān)基因的表達(dá),在自噬通路中發(fā)揮重要作用和功能,但具體作用機(jī)制目前尚不清楚。
2019年1月21日中國(guó)科學(xué)院生物物理研究所在《Autophagy》(IF:11.059)上在線(xiàn)發(fā)表了題為“YWHA/14-3-3 proteins recognize phosphorylated TFEB by a noncanonical mode for controlling TFEB cytoplasmic localization” 的文章揭示了YWHA/14-3-3與轉(zhuǎn)錄因子TFEB結(jié)合并調(diào)控TFEB亞細(xì)胞定位和轉(zhuǎn)錄活性的機(jī)制。

該研究中作者通過(guò)結(jié)構(gòu)生物學(xué)與生化手段,解析了YWHA/14-3-3與TFEB-p-S211復(fù)合物的結(jié)構(gòu),利用結(jié)構(gòu)信息發(fā)現(xiàn)YWHA/14-3-3與TFEB-p-S211以一種新的非典型模式結(jié)合。
通過(guò)進(jìn)一步對(duì)蛋白與小肽相互作用界面上的關(guān)鍵氨基酸進(jìn)行突變并結(jié)合細(xì)胞定位實(shí)驗(yàn),提出了潛在的YWHA/14-3-3 蛋白調(diào)節(jié) TFEB 的入核機(jī)制。這對(duì)于以 TFEB為靶點(diǎn)的新的治療方案的研究有指導(dǎo)作用。
作者首先對(duì)YWHA/14-3-3和TFEB兩個(gè)蛋白進(jìn)行二級(jí)結(jié)構(gòu)預(yù)測(cè)以及結(jié)構(gòu)域分析,結(jié)合已有的報(bào)道分析出磷酸化的TFEB中S142和S211兩個(gè)位點(diǎn)與調(diào)控TFEB的亞細(xì)胞定位有關(guān)(圖1. A,B)。
由吉?jiǎng)P基因構(gòu)建TFEB及其關(guān)鍵位點(diǎn)突變體的質(zhì)粒,通過(guò)免疫共沉淀實(shí)驗(yàn)(圖1. C)分別驗(yàn)證野生型TFEB 以及突變體與YWHA/14-3-3 蛋白兩種亞型的相互作用關(guān)系。通過(guò)ITC實(shí)驗(yàn)(圖1. D)比較它們之間的結(jié)合力大小,進(jìn)一步證明了只有磷酸化的TFEB中S211位點(diǎn)與YWHA/14-3-3 蛋白存在相互作用。

圖1. 磷酸化的TFEB中S211位點(diǎn)與YWHA/14-3-3 蛋白存在相互作用
作者分別解析了YWHA/14-3-3蛋白的兩種亞型YWHAB和YWHAG分別與TFEB-p-S211形成復(fù)合物的結(jié)構(gòu)。在復(fù)合物結(jié)構(gòu)中整體結(jié)構(gòu)為二聚體,在每個(gè)單體均由9個(gè)反向平行的 α 螺旋(αA-αI)構(gòu)成。αA-αD形成馬蹄鐵狀結(jié)構(gòu)域,并且二聚體的兩個(gè)亞基是雙面對(duì)稱(chēng)的(圖2. A-C)。
二聚體的每個(gè)亞基中,αC,αE,αG 和 αI 構(gòu)成一個(gè)兩親性的底物結(jié)合溝槽(活性中心),并特異性結(jié)合TFEB-p-S211 磷酸化小肽(圖2. D,E)。
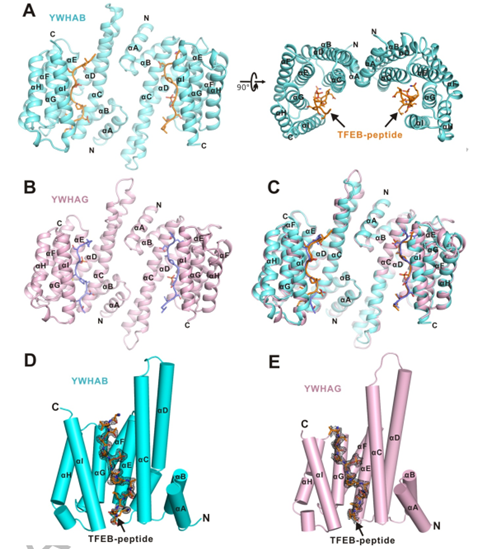
圖2. YWHA/14-3-3蛋白的兩種亞型分別與TFEB-p-S211復(fù)合物的整體結(jié)構(gòu)圖
作者選取了YWHAB 和 TFEB-p-S211復(fù)合物的結(jié)構(gòu)進(jìn)行后續(xù)詳細(xì)的結(jié)構(gòu)分析。YWHAB 和 TFEB-p-S211的結(jié)合主要是通過(guò)疏水作用力,靜電相互作用以及氫鍵相互作用。
小肽TFEB-p-S211頂部主要是通過(guò)S209側(cè)鏈以及E182與YWHAB形成氫鍵相互作用,中部通過(guò)R129、R58、Y130、K51以靜電荷和氫鍵相互作用協(xié)同幫助YWHAB 與 TFEB-p-S211結(jié)合,小肽TFEB-p-S211下部作用界面主要以疏水相互作用構(gòu)成(圖3)。
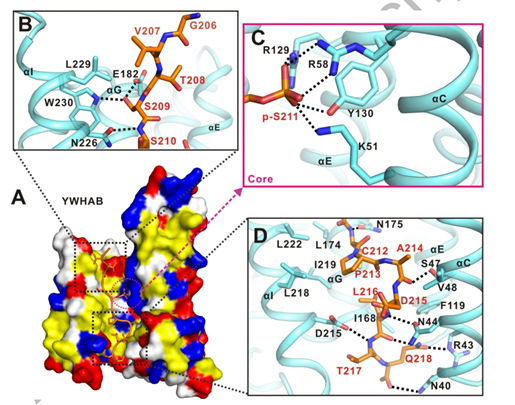
圖3. LECT2與Tie1直接結(jié)合
作者對(duì)現(xiàn)有的YWHA/14-3-3三種結(jié)合基序進(jìn)行了分析,已報(bào)道的YWHA/14-3-3與 mode I/II 磷酸化小肽復(fù)合物結(jié)構(gòu)中,-3 或-4 位的精氨酸在頂部結(jié)合區(qū)域,與YWHA/14-3-3的底物結(jié)合凹槽形成靜電相互作用。
mode III結(jié)合模式是識(shí)別羧基端末尾的磷酸化小肽,N 端的精氨酸消失。而文中提及的TFEB-p-S211結(jié)合YWHA/14-3-3 是一種新型非經(jīng)典的結(jié)合方式,缺少 N 端的靜電作用,但是多出了 C 端的疏水作用和氫鍵相互作用。因此,作者把它定義為 mode IV(圖4)。
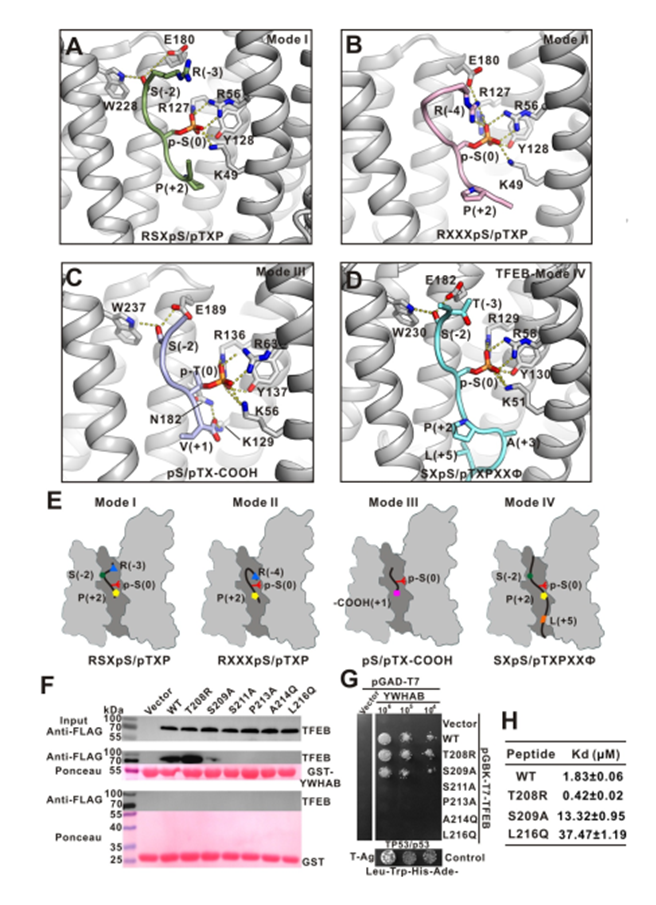
圖4. LECT2中斷Tie1 / Tie2異二聚化,Tie1對(duì)LECT2的功能行使發(fā)揮重要作用
作者通過(guò)對(duì)TFEB 全長(zhǎng)蛋白進(jìn)行了定點(diǎn)突變以破壞小肽與蛋白的相互作用界面,看是否能對(duì)TFEB 的亞細(xì)胞定位造成影響(圖5. A、B)。
實(shí)驗(yàn)表明靜息態(tài)的細(xì)胞在營(yíng)養(yǎng)充足的條件下,被mTORC1磷酸化的TFEB通過(guò)與銜接蛋白YWHA/14-3-3的相互作用被滯留在細(xì)胞質(zhì)中;而在細(xì)胞處于饑餓或者溶酶體功能障礙的狀態(tài)下,鈣調(diào)磷酸酶(calcineurin)使TFEB去磷酸化,導(dǎo)致TFEB與YWHA/14-3-3分離并進(jìn)入細(xì)胞核激活TFEB靶基因的轉(zhuǎn)錄。
因此YWHA/14-3-3是TFEB在細(xì)胞質(zhì)和細(xì)胞核之間位置轉(zhuǎn)換的關(guān)鍵調(diào)控因子。基于上述實(shí)驗(yàn)結(jié)果作者提出了潛在的 YWHA/14-3-3 蛋白調(diào)節(jié) TFEB 入核機(jī)制模型圖(圖5. C)。TFEB 的核定位序列與YWHA/14-3-3 結(jié)合位點(diǎn) S211空間距離很近,當(dāng) TFEB 和 YWHA/14-3-3蛋白結(jié)合可能會(huì)誘導(dǎo)pS211 和 NLS 之間的區(qū)域發(fā)生構(gòu)象變化,使 NLS 序列被遮擋從而導(dǎo)致不能細(xì)胞核錨定。
反之經(jīng)過(guò)饑餓處理后,S211位點(diǎn)被去磷酸化,TFEB 和 YWHA/14-3-3蛋白分離,NLS 序列暴露出來(lái),從而介導(dǎo)細(xì)胞核錨定和入核。
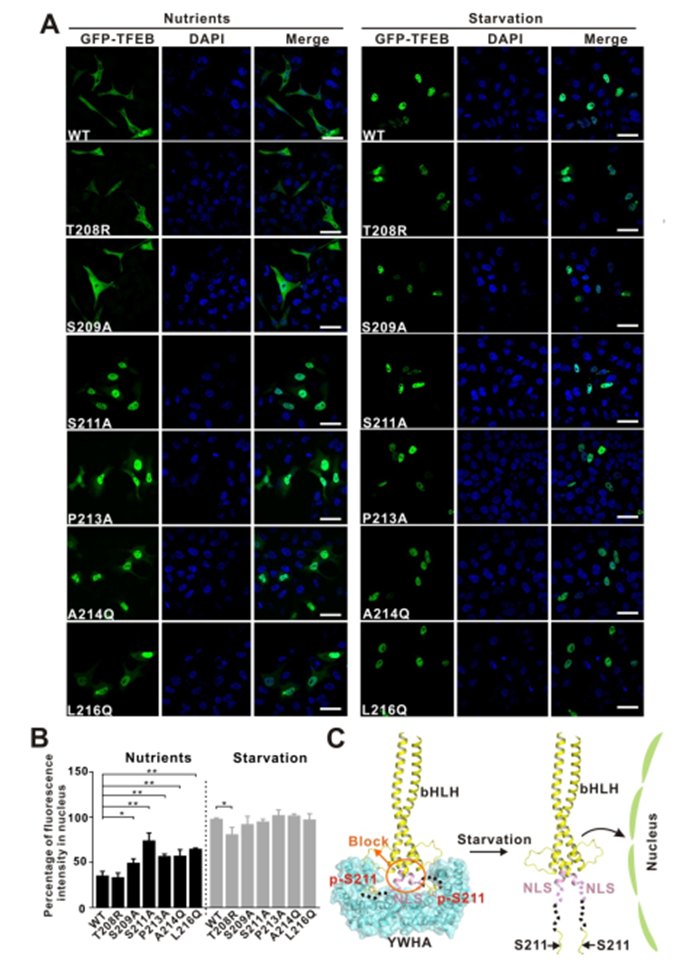
圖5. YWHA/14-3-3蛋白和 TFEB 之間的作用關(guān)系調(diào)控 TFEB 亞細(xì)胞定位
總結(jié)
1、 解析了YWHA/14-3-3蛋白的兩種亞型分別與TFEB-p-S211復(fù)合物的結(jié)構(gòu);
2、 提出YWHA/14-3-3蛋白通過(guò)非典型模式識(shí)別TFEB-p-S211;
3、 提出可能的 YWHA/14-3-3 蛋白調(diào)節(jié) TFEB 入核機(jī)制
吉?jiǎng)P助力
該研究的Co-IP實(shí)驗(yàn)部分,TFEB及其突變體的質(zhì)粒均由吉?jiǎng)P基因設(shè)計(jì)構(gòu)建,所用載體為pcDNA3.1-3×FLAG。其轉(zhuǎn)染HeLa細(xì)胞后有效的過(guò)表達(dá)TFEB及其突變體蛋白(圖6),進(jìn)而幫助證明了只有S211位點(diǎn)突變后會(huì)影響YWHA/14-3-3和TFEB的相互作用。

圖6. Western blot檢測(cè)TFEB及其突變體過(guò)表達(dá)效果
吉?jiǎng)P基因,年產(chǎn)上萬(wàn)次慢病毒、腺病毒、腺相關(guān)病毒;
建有國(guó)內(nèi)大型的慢病毒文庫(kù),包含幾乎覆蓋人類(lèi)所有基因的,近 15 萬(wàn)個(gè)獨(dú)立克隆;
國(guó)內(nèi)集產(chǎn)品采購(gòu)和資訊分享的一體化綜合服務(wù)平臺(tái),為科研工作者提供一鍵式的采購(gòu)服務(wù),產(chǎn)品周期短至 5 個(gè)工作日起;
17 年品質(zhì)保證、助力客戶(hù)發(fā)表10000+ 篇 SCI,多項(xiàng)科研成果發(fā)表于Nature Medicine、Nature Immunology、Gastroenterology、Cell Metabolism、Nature Neuroscience等頂級(jí)期刊上