
編者按
乳腺纖維腺瘤(FAs)是一種纖維上皮腫瘤,常見于青春期女性,長(zhǎng)期患有會(huì)增加患乳腺癌的風(fēng)險(xiǎn)。剖析人類乳腺FAs的病理機(jī)制對(duì)于開發(fā)更有效的治療方法至關(guān)重要。然而,凋亡的癌細(xì)胞對(duì)鄰近腫瘤上皮細(xì)胞的直接影響以及如何潛在促進(jìn)治療耐藥尚不清楚。
2023年6月18日,浙江大學(xué)醫(yī)學(xué)院附屬第二醫(yī)院、浙江省腫瘤微環(huán)境與免疫治療重點(diǎn)實(shí)驗(yàn)室黃建、陳志剛團(tuán)隊(duì)在《Nature Communications》雜志發(fā)表了題為“Single cell profiling of female breast fibroadenoma reveals distinct epithelial cell compositions and therapeutic targets”的研究論文。該研究利用高通量單細(xì)胞RNA-seq分析,發(fā)現(xiàn)了乳腺纖維腺瘤中不同的細(xì)胞組成和上皮結(jié)構(gòu)變化,證實(shí)上皮細(xì)胞可表現(xiàn)出激素響應(yīng)功能特征,并同步激活雌激素敏感和激素抗性機(jī)制。此外,研究人員還建立了一個(gè)可擴(kuò)展的患者來源的FA類器官平臺(tái),通過對(duì)通路抑制劑進(jìn)行系統(tǒng)探究以測(cè)試FAs的治療反應(yīng),以期為乳腺FAs提供潛在治療策略。
原文獻(xiàn):Chen, Z., Zhang, Y., Li, W. et al. Single cell profiling of female breast fibroadenoma reveals distinct epithelial cell compositions and therapeutic targets. Nat Commun 14, 3469 (2023).
一、 研究背景
乳腺纖維腺瘤(FAs)是一種纖維上皮腫瘤,常見于青春期女性,長(zhǎng)期患有會(huì)增加患乳腺癌的風(fēng)險(xiǎn)[1]。手術(shù)切除是FAs最有效的治療措施,但可能會(huì)形成不良疤痕或廣泛的導(dǎo)管損傷[2]。由于這類患者往往有多個(gè)病灶部位,依靠手術(shù)難以全部根除,因此迫切需要非手術(shù)的治療措施。
FAs在臨床上被認(rèn)為具有激素依賴性[3]。然而,在以往的臨床試驗(yàn)中,雌激素受體調(diào)節(jié)劑他莫昔芬的治療效果并不理想[4, 5]。因此,剖析人類乳腺FAs的病理機(jī)制對(duì)于開發(fā)更有效的治療方法至關(guān)重要。盡管大量測(cè)序研究已經(jīng)揭示高復(fù)發(fā)FAs的突變基因并繪制出相關(guān)基因組圖譜,但由于疾病本身的復(fù)雜性以及缺乏可重復(fù)的人體模型,F(xiàn)As和正常乳腺組織之間上皮細(xì)胞和基質(zhì)細(xì)胞轉(zhuǎn)錄差異的潛在機(jī)制在很大程度上仍然未知。
本研究利用高通量單細(xì)胞RNA-seq分析,發(fā)現(xiàn)了FAs不同的細(xì)胞組成和上皮結(jié)構(gòu)變化,證實(shí)上皮細(xì)胞可表現(xiàn)出激素響應(yīng)功能特征,并同步激活雌激素敏感和激素抗性機(jī)制。此外,研究人員還建立了一個(gè)可擴(kuò)展的患者來源的FA類器官平臺(tái),通過對(duì)通路抑制劑進(jìn)行系統(tǒng)探究以測(cè)試FAs的治療反應(yīng),以期為乳腺FAs提供潛在治療策略。
二、 研究結(jié)果
1、單細(xì)胞轉(zhuǎn)錄組分析確定了FAs的細(xì)胞組成和上皮結(jié)構(gòu)特征
對(duì)4個(gè)乳腺FAs和2個(gè)配對(duì)的癌旁乳腺組織進(jìn)行單細(xì)胞RNA測(cè)序(圖1a),根據(jù)不同的基因表達(dá)特征,這些細(xì)胞被劃分為10個(gè)細(xì)胞簇:成熟的管腔細(xì)胞(ML)、管腔祖細(xì)胞(LP)和基底細(xì)胞(BC)、成纖維細(xì)胞(FB)、內(nèi)皮細(xì)胞(EC)、周細(xì)胞(PT)、T細(xì)胞(T)、B細(xì)胞(B)、骨髓細(xì)胞(M)以及未知細(xì)胞(Un)(圖1b)。
與正常乳腺組織相比,F(xiàn)As中的基質(zhì)細(xì)胞幾乎不表達(dá)激素受體,而上皮細(xì)胞則表達(dá)了高水平的激素受體。免疫組織化學(xué)染色結(jié)果證實(shí)激素受體主要在上皮細(xì)胞而非基質(zhì)細(xì)胞中表達(dá)(圖2d)。成纖維細(xì)胞在FAs在基質(zhì)細(xì)胞中顯著增多(圖1c),但增殖基因(MKI67、PCNA)和KI67免疫組化染色的分析顯示,成纖維細(xì)胞的增加可能不是由更高的增殖引起的。研究人員從腫瘤組織中分離出原代上皮細(xì)胞和成纖維細(xì)胞,并配對(duì)正常組織,進(jìn)行上皮細(xì)胞和成纖維細(xì)胞的共培養(yǎng)。結(jié)果顯示正常乳腺組織中的成纖維細(xì)胞可以被FA上皮募集,證明上皮細(xì)胞在FAs形成中的重要作用。
scRNA數(shù)據(jù)顯示,與正常乳腺上皮細(xì)胞相比,F(xiàn)As的上皮細(xì)胞組成中成熟管腔細(xì)胞擴(kuò)大,基底細(xì)胞減少(圖1c)。研究人員基于EpCAM和CD49f的表達(dá)組合對(duì)59例新鮮FA組織及組織和21例正常乳腺組織的上皮亞群進(jìn)行表征,證實(shí)了這一現(xiàn)象。已知正常乳腺由外層基底細(xì)胞和內(nèi)腔細(xì)胞組成[6]。FAs中基底細(xì)胞的損失表明雙層結(jié)構(gòu)發(fā)生了變化。這些數(shù)據(jù)表明FAs的細(xì)胞組成和結(jié)構(gòu)特征與正常乳腺上皮細(xì)胞不同。
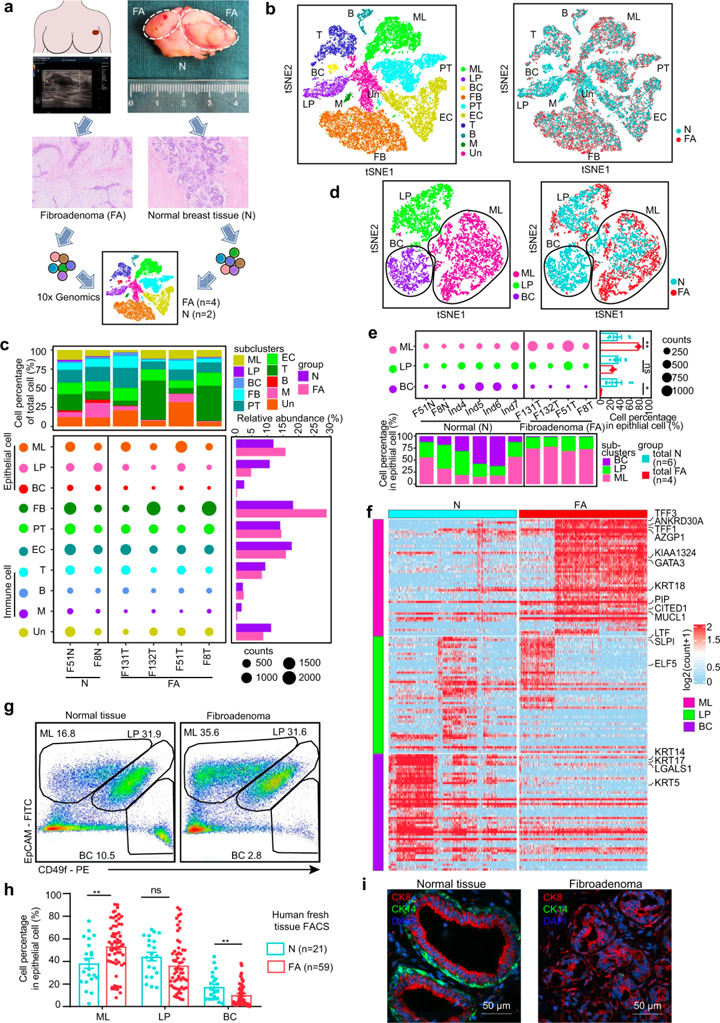
圖1
2、FAs中的上皮細(xì)胞顯示激素反應(yīng)性特征
接著,研究人員對(duì)FAs上皮細(xì)胞群的功能差異進(jìn)行探究。GO分析顯示FAs上皮細(xì)胞與激素反應(yīng)密切相關(guān)(圖2a),基因富集顯示受雌激素和雌激素誘導(dǎo)基因調(diào)節(jié)的TFF1和TFF3變化最為顯著(圖2c)。
免疫熒光和免疫組織化學(xué)染色進(jìn)一步證實(shí)了FAs患者中這些激素相關(guān)蛋白的表達(dá)模式(圖2d,e)。對(duì)3種不同上皮細(xì)胞類型的DEGs進(jìn)行比較:(1)成熟管腔細(xì)胞上調(diào)的基因主要與激素反應(yīng)信號(hào)通路相關(guān),且管腔細(xì)胞能夠響應(yīng)內(nèi)分泌信號(hào)或產(chǎn)生乳汁[7];(2)管腔祖細(xì)胞上調(diào)的基因主要與上皮細(xì)胞增殖相關(guān);(3)基底細(xì)胞上調(diào)的基因主要與腺體發(fā)育的調(diào)節(jié)相關(guān)。這些結(jié)果表明FAs中的上皮細(xì)胞具有激素相應(yīng)的功能特征。
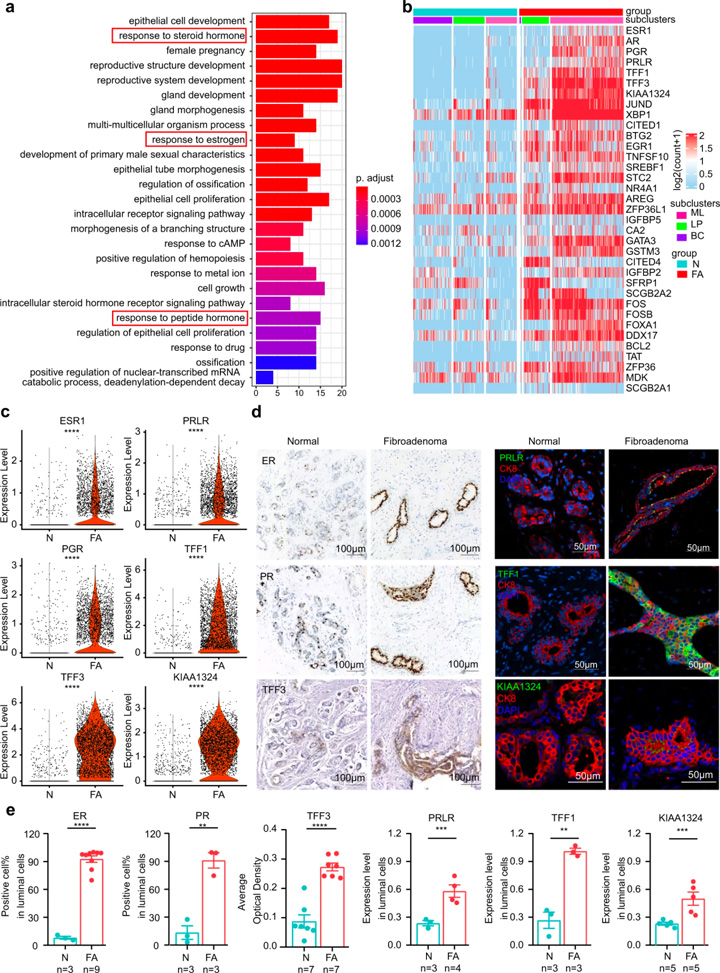
圖2
3、FAs中激素敏感性和激素抵抗性共存
臨床研究顯示激素敏感性通路可以控制FAs[3, 8]。研究人員通過KEGG分析發(fā)現(xiàn),除了激素敏感通路外,內(nèi)分泌抵抗通路也參與其中,GSEA也提供了類似的結(jié)果(圖3a, b)。參與內(nèi)分泌抵抗性通路的基因主要分為3個(gè)功能類別:ERBB2/IGF1R、CCND1和BCL2(圖3c),據(jù)報(bào)道這些基因與內(nèi)分泌抵抗[9-11]及乳腺癌復(fù)發(fā)風(fēng)險(xiǎn)增加有關(guān)[12, 13]。回顧15例術(shù)后3年內(nèi)同一乳房部位復(fù)發(fā)的FAs患者和9例無復(fù)發(fā)患者的資料,結(jié)果顯示復(fù)發(fā)FAs組中ERBB2、CCND1和BCL2的表達(dá)水平顯著高于未復(fù)發(fā)組(圖3d),且與FAs復(fù)發(fā)呈正相關(guān)。此外,乳腺癌患者的ERBB2、CCND1和BCL2表達(dá)水平顯著高于FAs(圖3e)。
對(duì)內(nèi)分泌抗性通路標(biāo)志物是否存在于每個(gè)患者的上皮細(xì)胞進(jìn)行探究。tSNE分析顯示,每位患者的上皮細(xì)胞中均能檢測(cè)到ERBB2+細(xì)胞、BCL2+細(xì)胞和CCND1+細(xì)胞。對(duì)4名FAs患者進(jìn)行免疫染色,結(jié)合單細(xì)胞RNA測(cè)序以進(jìn)一步驗(yàn)證這些發(fā)現(xiàn)。結(jié)果在正常人體組織中表達(dá)水平較低的激素敏感標(biāo)記物(ER、TFF1、TFF3)和內(nèi)分泌抵抗標(biāo)記物(ERBB2、BCL2、CCND1)在FAs患者中均被檢出,且樣本間具有顯著的異質(zhì)性,提示每個(gè)受試者對(duì)內(nèi)分泌治療具有不同的敏感性。這些數(shù)據(jù)表明FAs中激素敏感性和激素抵抗性共存。
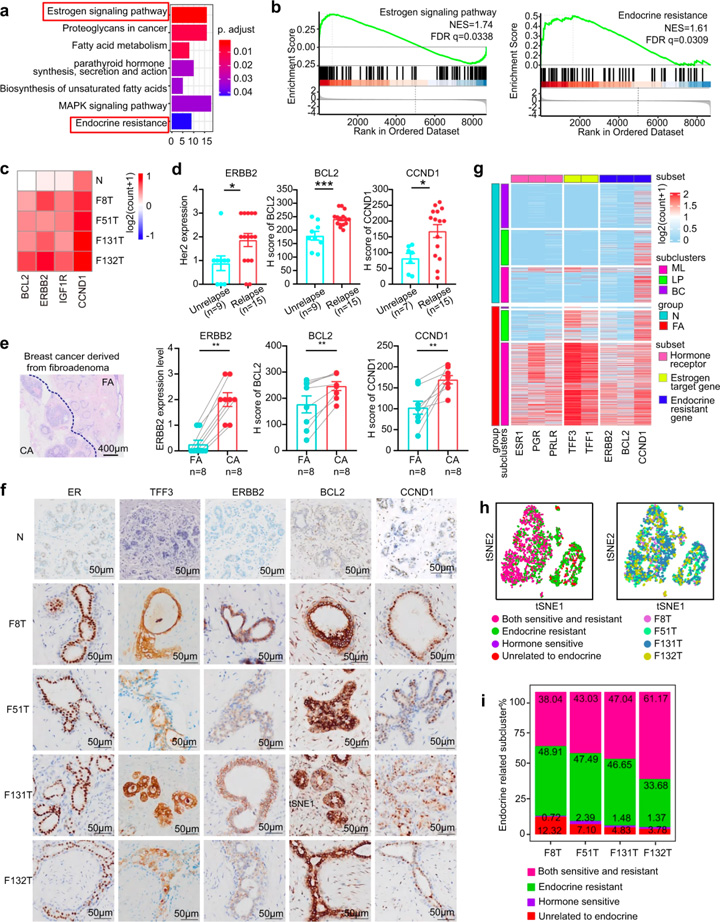
圖3
4、他莫昔芬與CCND1、BCL2或ERBB2抑制劑的聯(lián)合用藥可顯著抑制了他莫昔芬耐藥FAs類器官的活力
由于激素敏感和激素耐藥途徑在FAs中共存,研究人員探索了為FAs患者提供個(gè)體化內(nèi)分泌治療的可能性。對(duì)39例FA患者開發(fā)了一個(gè)可擴(kuò)展的類器官平臺(tái)。這些類器官的組織學(xué)特征與原始FA上皮非常相似(圖4b),全外顯子組測(cè)序結(jié)果顯示類器官總體上保留了人類纖維腺瘤的基因組結(jié)構(gòu)(圖4c)。RNA測(cè)序顯示,相對(duì)于正常上皮,類器官和FAs之間激素敏感和激素抗性標(biāo)記的轉(zhuǎn)錄變化相似(圖4d)。
與正常乳腺類器官相比,F(xiàn)As類器官中激素敏感標(biāo)記物和耐藥通路標(biāo)記物也上調(diào)(圖4d)。GSEA還揭示了人類FAs類器官中激素敏感和激素抗性通路的陽性富集(圖4e)。FACS進(jìn)一步證明,人FAs衍生類器官的上皮細(xì)胞與原始FAs具有幾乎相同的細(xì)胞組成(圖4f)。為了驗(yàn)證激素響應(yīng)功能特征,用10 nM雌二醇處理FA衍生的類器官,顯示ER響應(yīng)的標(biāo)志基因顯著上調(diào),且刺激了類器官的生長(zhǎng)。這些數(shù)據(jù)表明FA衍生的類器官是激素反應(yīng)性的,可作為人類FAs的模型代表,用于開發(fā)個(gè)體化內(nèi)分泌治療。
13例類器官的藥敏結(jié)果顯示不同患者對(duì)他莫昔芬的反應(yīng)存在異質(zhì)性(圖4g)。大多數(shù)類器官的IC50值要高于臨床他莫昔芬耐藥患者。基于不同人FA衍生的類器官中他莫昔芬耐藥途徑標(biāo)志物的差異表達(dá),將他莫昔芬與CDK4/6抑制劑Palbociclib、BCL2抑制劑Venetoclax和ERBB2抑制劑lapatinib聯(lián)合,在IC10的濃度下進(jìn)行藥物敏感性篩選(圖4h-j)。結(jié)果顯示他莫昔芬與CCND1、BCL2或ERBB2抑制劑的用藥組合顯著抑制了他莫昔芬耐藥FAs的生存能力,為FAs的干預(yù)提供了潛在治療方法。
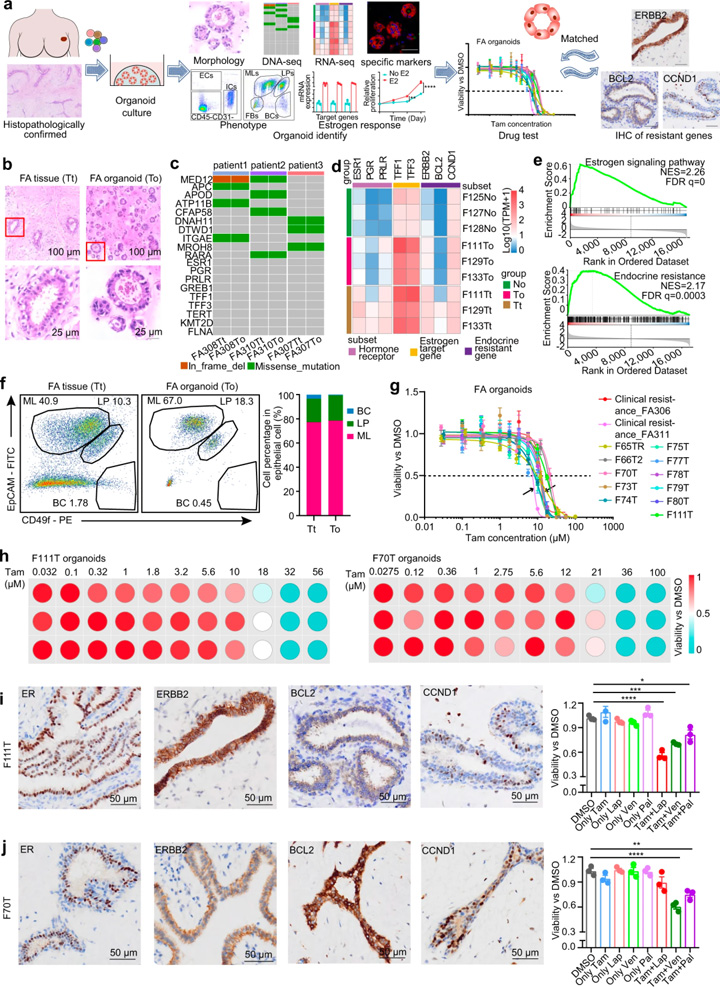
圖4
作為全球斑馬魚技術(shù)應(yīng)用的領(lǐng)導(dǎo)者,環(huán)特生物構(gòu)建了“斑馬魚、類器官、哺乳動(dòng)物、人體”四位一體的綜合技術(shù)服務(wù)平臺(tái),開展科研及研究服務(wù)、智慧實(shí)驗(yàn)室建設(shè)和精準(zhǔn)醫(yī)療三大業(yè)務(wù)。目前,環(huán)特類器官平臺(tái)已成功搭建多種腫瘤及正常類器官的培養(yǎng)平臺(tái),已有多種腫瘤類器官培養(yǎng)試劑盒和試劑在售,歡迎各位讀者垂詢!
參考文獻(xiàn)
[1] Dupont, W. D. et al. Long-term risk of breast cancer in women with fibroadenoma. N. Engl. J. Med. 331, 10–15 (1994).
[2]. Lee, M. & Soltanian, H. T. Breast fibroadenomas in adolescents: current perspectives. Adolesc. Health Med. Ther. 6, 159–163 (2015).
[3]. Yu, J. H. et al. Breast diseases during pregnancy and lactation. Obstet. Gynecol. Sci. 56, 143–159 (2013).
[4]. Viviani, R. S., Gebrim, L. H., Baracat, E. C. & De Lima, G. R. Evaluationof the ultrasonographic volume of breast fibroadenomas in womentreated with tamoxifen. Minerva Ginecol. 54, 531–535 (2002).
[5]. Bernardes, J. R. Jr., Seixas, M. T., Lima, G. R., Marinho, L. C. & Gebrim, L. H. The effect of tamoxifen on PCNA expression in fibroadenomas. Breast J. 9, 302–306 (2003)
[6]. Fu, N. Y., Nolan, E., Lindeman, G. J. & Visvader, J. E. Stem cells and the differentiation hierarchy in mammary gland development. Physiol. Rev. 100, 489–523 (2020).
[7]. Li, C. M. et al. Aging-associated alterations in mammary epithelia and stroma revealed by single-cell RNA sequencing. Cell Rep. 33, 108566 (2020).
[8]. Greenberg, R., Skornick, Y. & Kaplan, O. Management of breast fibroadenomas. J. Gen. Intern. Med. 13, 640–645 (1998).
[9]. Hanker, A. B., Sudhan, D. R. & Arteaga, C. L. Overcoming endocrine resistance in breast cancer. Cancer Cell 37, 496–513 (2020).
[10]. Choi, H. J. et al. Role of RBP2-induced ER and IGF1R-ErbB signaling in tamoxifen resistance in breast cancer. J. Natl Cancer Inst. 110, https://doi.org/10.1093/jnci/djx207 (2018
[11]. Rozeboom, B., Dey, N. & De, P. ER+ metastatic breast cancer: past, present, and a prescription for an apoptosis-targeted future. Am. J. Cancer Res. 9, 2821–2831 (2019).
[12]. Slamon, D. J. et al. Human breast cancer: correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 235, 177–182 (1987).
[13]. Roy, P. G. et al. High CCND1 amplification identifies a group of poor prognosis women with estrogen receptor positive breast cancer. Int. J. Cancer 127, 355–360 (2010).