大腦對意識的加工依賴于大腦皮層內的活動[1]。其中小鼠皮層可被分為六層,每層都由不同的神經元細胞類型組成,每一層的細胞都參與皮層微環路的形成。每一層內的興奮性細胞可接受來自同一層內其他興奮性細胞和抑制細胞的輸入[2]。其中L5中的興奮性錐體神經元(pyramidal neurons,PNs)根據其輸出的不同分為三種類型:向其他皮層區域提供遠程輸出的IT(intratelencephalic)神經元;連接TH和其他皮層下區域的錐體束(PT)神經元和與其他L5 PNs連接的NP(near-projecting)神經元[3]。
所有全身性***均可導致意識喪失[4]。臨床上常見的***包括異氟烷(Isoflurane,Iso)、芬太尼-美托咪定-咪達**(Fentanyl-Medetomidine-Midazolam, FMM)和***-甲苯噻嗪(Ketamine-Xylazine ,Ket-Xyl);雖然這幾種***介導麻醉的分子機制不盡相同,但是麻醉誘導的無意識行為和皮層腦電活動的變化是相同的,并在意識喪失時出現,在意識恢復時逆轉[5]。除了腦電的變化,fMRI也觀察到無意識狀態下皮質-皮質和皮質-丘腦聯系的斷開。在全身麻醉期間,包括L5和L6神經元在內的電特性為長程投射神經元的神經元表現出與腦電圖和ECoG觀察到的頻率變化相關的活動變化[5]。
因為不同類型的神經元在控制皮層微環路中的信息傳遞有不同的功能,測量單個細胞類型的活動可以更好的了解無意識狀態下神經元電活動的相關性。但對麻醉情況下不同神經元的電活動的了解還了解甚少。
最近,來自瑞士巴塞爾分子與眼科研究所的Botond Roska課題組在Neuron發表了題為General anesthesia globally synchronizes activity selectively in layer 5 cortical pyramidal neurons的文章,發現了全身***持續影響小鼠L5椎體神經元自發電活動但不影響其他細胞類型電活動的機制[6]。

01.麻醉引起L5 PNs的同步活動
為了特異性觀察L5 PNs的電活動,研究者們使用Rbp4-Cre KL100小鼠,并通過AAV-CAG-FLEX-GCaMP7s記錄鈣成像。研究者們分別用FMM、Iso和Ket-Xyl三種不同的條件誘導小鼠麻醉,結果發現不論在清醒狀態還是麻醉狀態,L5 PNs都表現出自發的鈣活動(圖1C),但與清醒時相比,麻醉狀態下L5神經元鈣活動的同步性更高(圖1D)。盡管在麻醉狀態下L5神經元放電頻率和振幅也發生了變化,但三種***引起的變化不大一致(圖2A-D)。
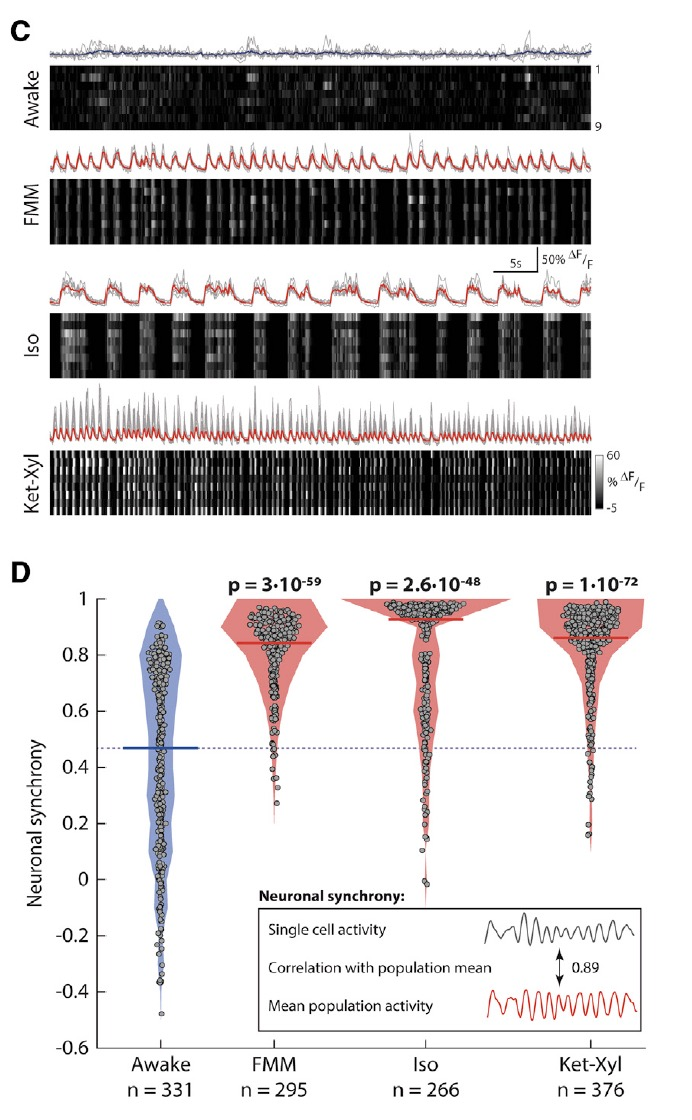
圖1:麻醉狀態下L5神經元電活動同步性更高
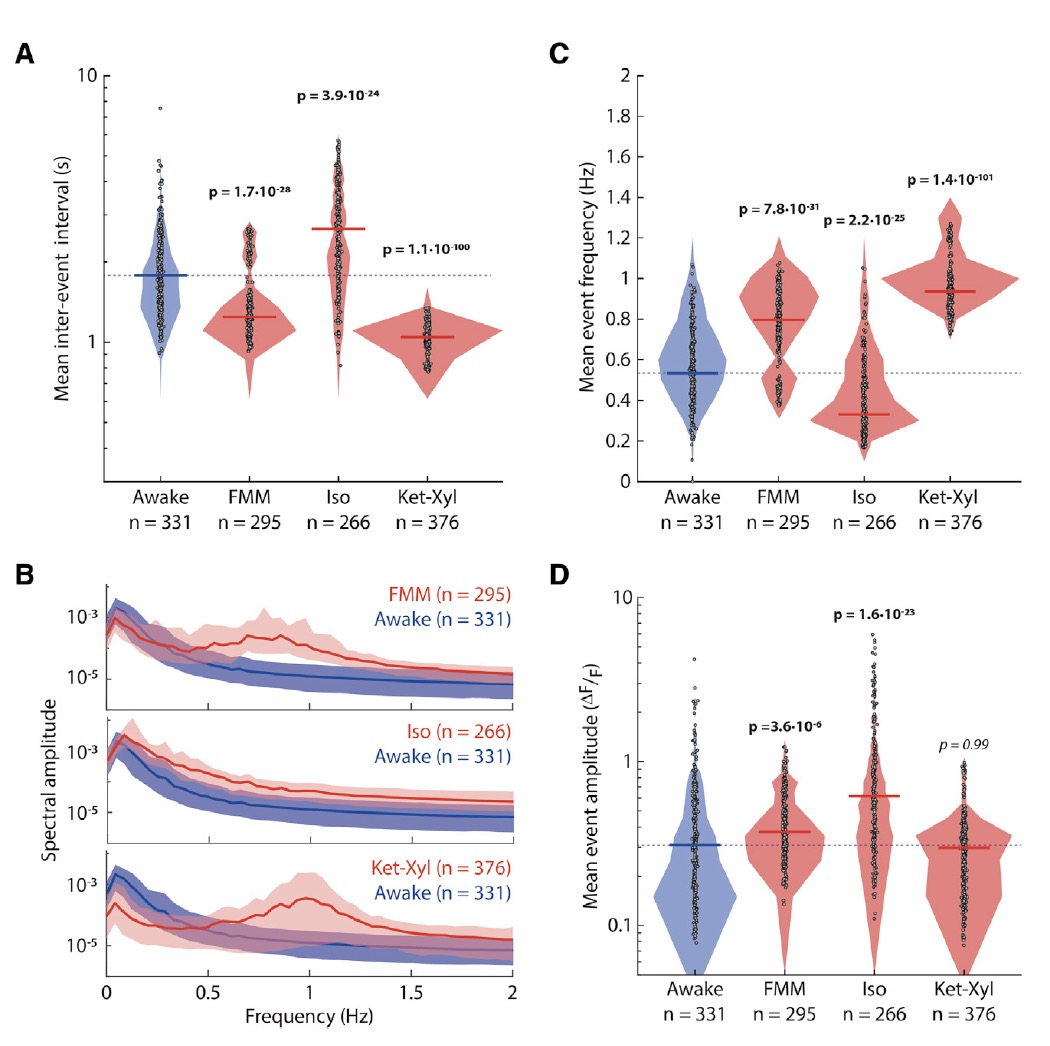
圖2:麻醉狀態下L5神經元電活動頻率和幅度的變化
02. L5 PNs同步性與意識的喪失恢復有關
接下來研究者們想弄清楚L5 PNs同步性增高的生理意義,于是他們將電活動同步性與小鼠行為(眼睛轉動、胡須抖動和身體運動)進行了關聯分析。研究結果顯示,神經元同步性的轉化與小鼠運動行為的轉化相關性非常好(R=0.89),統計學分析也證明神經元同步性的轉變時間與運動行為的轉變時間無顯著差異(圖3D、H)。
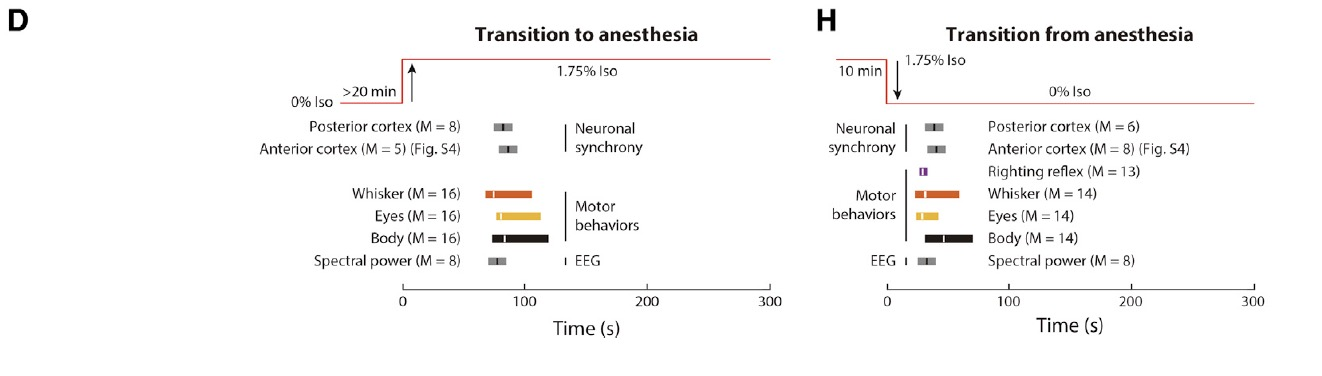
圖3:麻醉狀態下L5神經元同步性與意識變化相關
03. 神經元同步性增強僅發生在L5 PNs
那么這種神經元同步性的增強是否發生在其他類型的神經元中?接下來研究者們對初級視覺皮層L1,L2/3,L4,L6麻醉后的神經元同步性進行了分析(圖4A)。結果發現對于所有被研究的細胞類型,三種麻藥引起的麻醉狀態下神經元的同步性并不一致(圖4B-F)。這說明麻醉引起的神經元同步性變強是L5 PNs特異的。
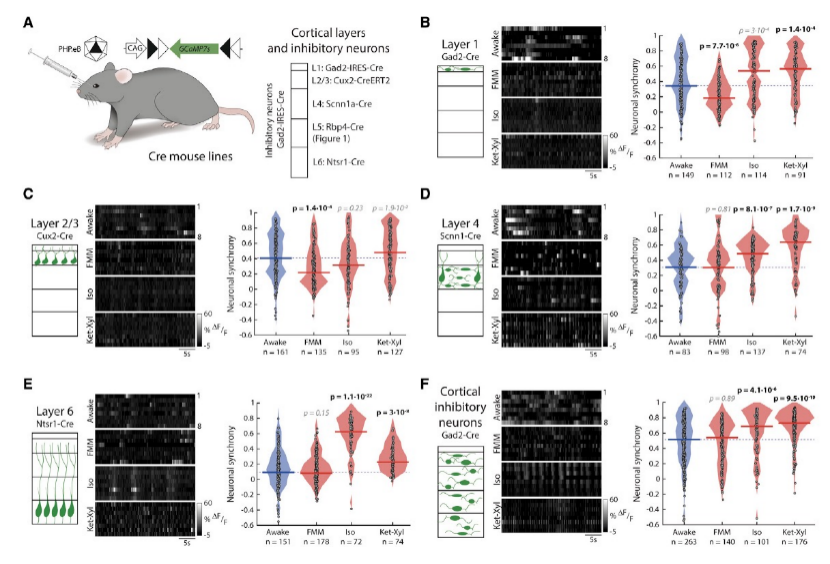
04. 麻醉時投射至L5 的突觸前神經元活動被抑制
為了進一步研究L5 PNs同步性變化的機制,研究者們對L5神經元的輸入進行了研究。他們采用PHP.eB AAV-syn-FLEX-splitTVA-EGFP-tTA ,PHP.eB AAV-TREtight-mTagBFP2-B19G 和EnvA-SADDG-GCaMP7s rabies virus聯用的雙AAV策略實現單突觸逆行跨突觸的標記(圖5A),在感染后5天可觀察到L5外的突觸前神經元成功表達GCaMP7s(圖B)。分別對清醒和麻醉狀態下神經元電活動記錄發現,這些突觸前神經元在清醒時是活躍的,但Iso麻醉引起突觸前神經元的活動顯著降低(圖5C-E)。
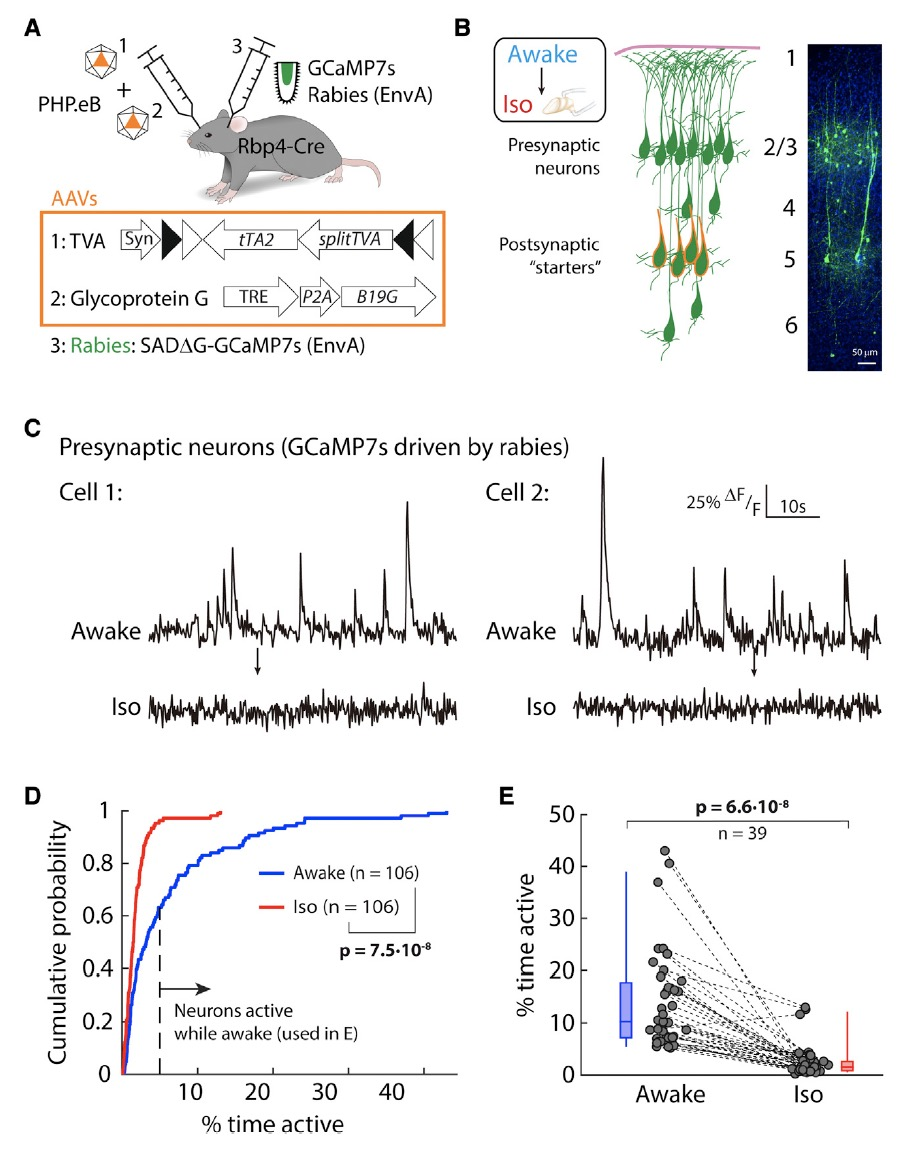
圖5: L5突觸前神經元電活動在麻醉時顯著降低
05. L5 PNs在皮質區域的活動是同步的
由于之前的研究只是針對視覺皮層L5進行了成像,為了對整個皮質L5 PNs進行研究,研究者們向Rbp4-Cre小鼠注射AAV-CAG-FLEX-GCaMP7s,并對皮層的一個大區域進行了廣域的熒光成像(圖6A)。通過對多個撓區清醒和麻醉狀態下鈣活動的記錄,他們發現麻醉的確會導致皮層廣泛區域的L5 PNs神經元同步性增高(圖6C)。
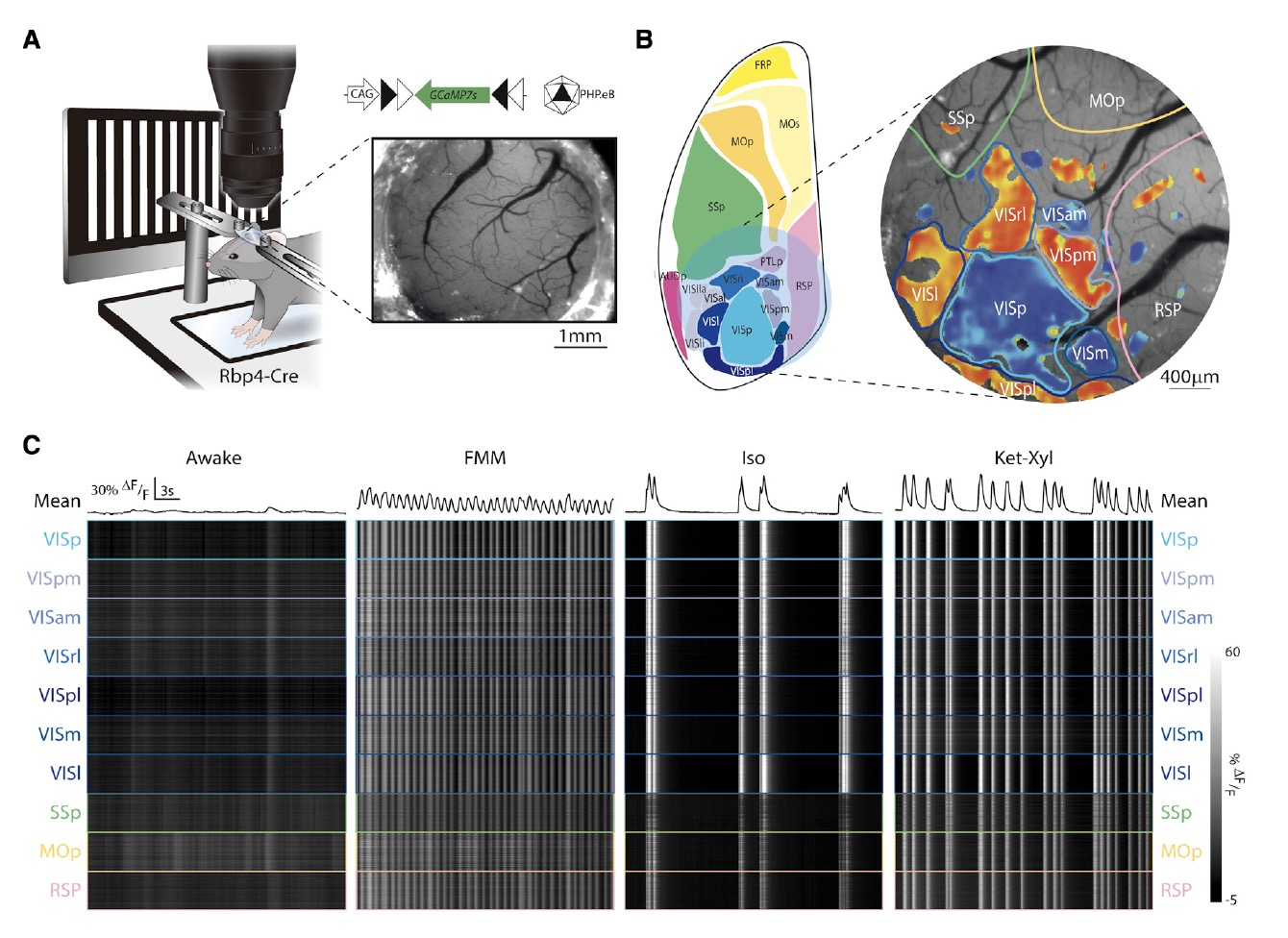
圖6: L5同步電活動出現在多個皮層區域
總之,這項研究揭示了了小鼠L5錐體神經元電活動與麻醉的關系,發現了不同***物引起L5 PNs同步性增強的現象,并且這種高度的神經元同步性在皮層大范圍存在。且在進入麻醉狀態和從麻醉狀態中復蘇的過程中,L5 PNs的變化與意識的喪失和恢復相一致。
本文利用鈣離子成像等多種AAV,成功研究了小鼠在清醒/麻醉狀態下L5 PNs 的電活動。
吉凱基因可為您提供多種AAV產品
定制AAV:過表達/干擾、CRISPR/Cas9(MIT/哈佛大學**授權)、Cre-on/off系統、特異性表達系統。滴度可達1.0E+13 v.g./mL以上
現貨AAV :鈣離子成像;光遺傳學;化學遺傳學;重組酶;熒光對照;順行逆行神經環路示蹤病毒
體內精準靶向研究利器——組織特異性AAV
40+的組織特異性啟動子,靶向神經/心血管/肝臟/骨/脂肪/肌肉/視網膜等,實現目的基因體內特異性表達

參考文獻…
向上滑動閱覽
[1] Brown, R., Lau, H., and LeDoux, J.E. (2019). Understanding the higher-order approach to consciousness. Trends Cogn. Sci. 23, 754–768.
[2] Koch, C., Massimini, M., Boly, M., and Tononi, G. (2016). Neural correlates of consciousness: progress and problems. Nat. Rev. Neurosci. 17, 307–321.
[3] Kim, E.J., Juavinett, A.L., Kyubwa, E.M., Jacobs, M.W., and Callaway, E.M. (2015). Three types of cortical Layer 5 neurons that differ in brain-wide connectivity and function. Neuron 88, 1253–1267.
[4] Hudetz, A.G., and Mashour, G.A. (2016). Disconnecting consciousness: is there a common anesthetic end-point? Anesth. Analg. 123, 1228–1240.
[5] Hagihira, S. (2015). Changes in the electroencephalogram during anaesthesia and their physiological basis. Br. J. Anaesth. 115, i27–i31.
[6] Bharioke et al., General anesthesia globally synchronizes activity selectively in layer 5 cortical pyramidal neurons, Neuron (2022), https://doi.org/10.1016/j.neuron.2022.03.032