代謝綜合征與心血管疾病相關,一定程度上是因為引起了動脈硬化。高脂高糖飲食是代謝綜合征的重要誘發因素,當前,他汀類降脂藥對于動脈硬化的改善效果并不理想。因此,闡明代謝綜合征導致動脈硬化的分子機制對于減少或預防后續的心血管相關疾病有重要意義。
今年,天津醫科大學基礎醫學院艾玎教授課題組在Circultion Research(中科院JCR一區,IF:23.213 )上發表了題為“YAP Targets the TGFβ Pathway to Mediate High-Fat/High-Sucrose Diet-Induced Arterial Stiffness”的文章,揭示了平滑肌細胞YAP分子在高脂高糖飲食誘導的動脈硬化中的作用和分子機制,為相關心血管疾病的治療提供了新的思路和靶點。
研究結果
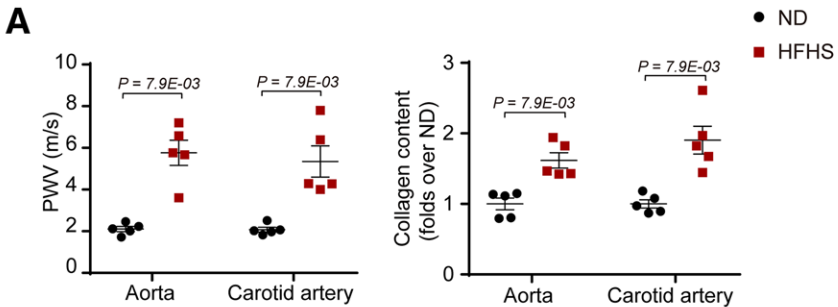
1. 高脂高糖飲食誘導小鼠動脈硬化
研究者將8周齡雄性小鼠隨機分為兩組,并分別喂食高脂高糖食物(HFHS)和正常食物(ND),8周后檢測動脈血管變化。與ND組相比,HFHS組小鼠主動脈和頸動脈的脈搏波速度和膠原含量均顯著上升,而血壓在兩組間相似。TGFβ已被證明是組織纖維化和細胞外基質產生的最重要的調控因子之一,因此,研究者檢測了TGFβ/Smad信號通路的激活。檢測結果表明,與ND小鼠相比,磷酸化Smad2/3的水平在HFHS組小鼠的主動脈中膜中顯著升高。
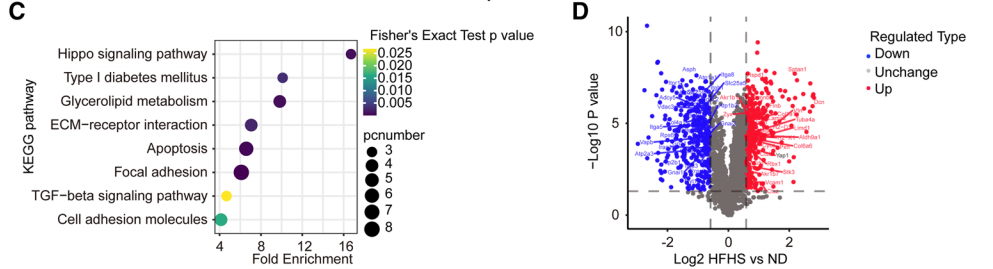
2. 4D蛋白質組學篩選關鍵蛋白
為了進一步揭示高脂高糖飲食誘導的TGFβ通路激活的機制,研究者對小鼠主動脈中膜(主要由血管平滑肌細胞(VSMC)組成,HFHS(n=3)vs. ND(n=3))進行4D蛋白質組學檢測。4D蛋白質組學共定量了2543個蛋白,其中341個上調蛋白,343個下調蛋白(fc>1.5,p<0.05)。差異上調蛋白的KEGG分析顯示了ECM受體相互作用和TGFβ信號通路及膽固醇和甘油脂代謝途徑相關通路在HFHG小鼠主動脈中的富集。該結果表明已構建的小鼠模型與代謝紊亂誘導的動脈硬化相一致。研究者注意到,差異上調蛋白中存在一個與動脈硬化相關的蛋白——YAP。
研究者對YAP蛋白的表達情況進行驗證,發現在HFHS組小鼠中,YAP蛋白總體表達量上調,同時細胞核和細胞質中的YAP水平均上升,而YAP的細胞核比細胞質的比率降低。該結果意味著,血管平滑肌細胞(VSMC)中YAP蛋白的高表達在高脂高糖誘導的動脈硬化中有重要作用。
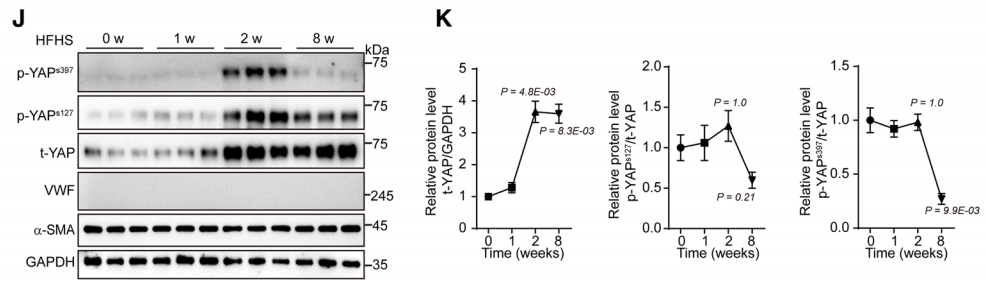
3. YAP蛋白隨時間的變化
研究者對高脂高糖喂食小鼠不同時間點YAP蛋白的變化進行檢測,發現在早期(第二周)YAP蛋白的表達量即呈現明顯的上調,磷酸化YAP在第二周出現明顯上調,而在第八周則下調。研究者檢測了第二周時YAP mRNA的表達水平,發現HFHS組和ND組的YAP mRNA表達量相當。這意味著,YAP的上調并不發生在轉錄水平上,而是在蛋白水平上。此外,HFHS組的總YAP和磷酸化YAP水平均顯著高于ND組。
考慮到mTORC1在過量營養攝入時會被激活,研究者的結果也表明在HFHS組中mTORC1活性增強。研究者在人動脈血管平滑肌細胞中使用siRNA抑制mTORC1,發現當mTORC1被抑制后,誘導了細胞自噬和YAP蛋白表達的下調,而這種作用在自噬抑制劑處理后得以恢復。此外,使用mTORC1抑制劑處理也會顯著降低YAP蛋白表達。綜合這些結果,研究者認為,高脂高糖飲食誘導的mTORC1激活導致的YAP表達上調是通過降低主動脈中膜的細胞自噬實現的。
有趣的是,脂肪、組織、肝和心臟組織中YAP的mRNA和蛋白水平并未受到高糖高脂飲食的影響,這意味著高脂高糖特異性誘導了血管中YAP蛋白的上調。
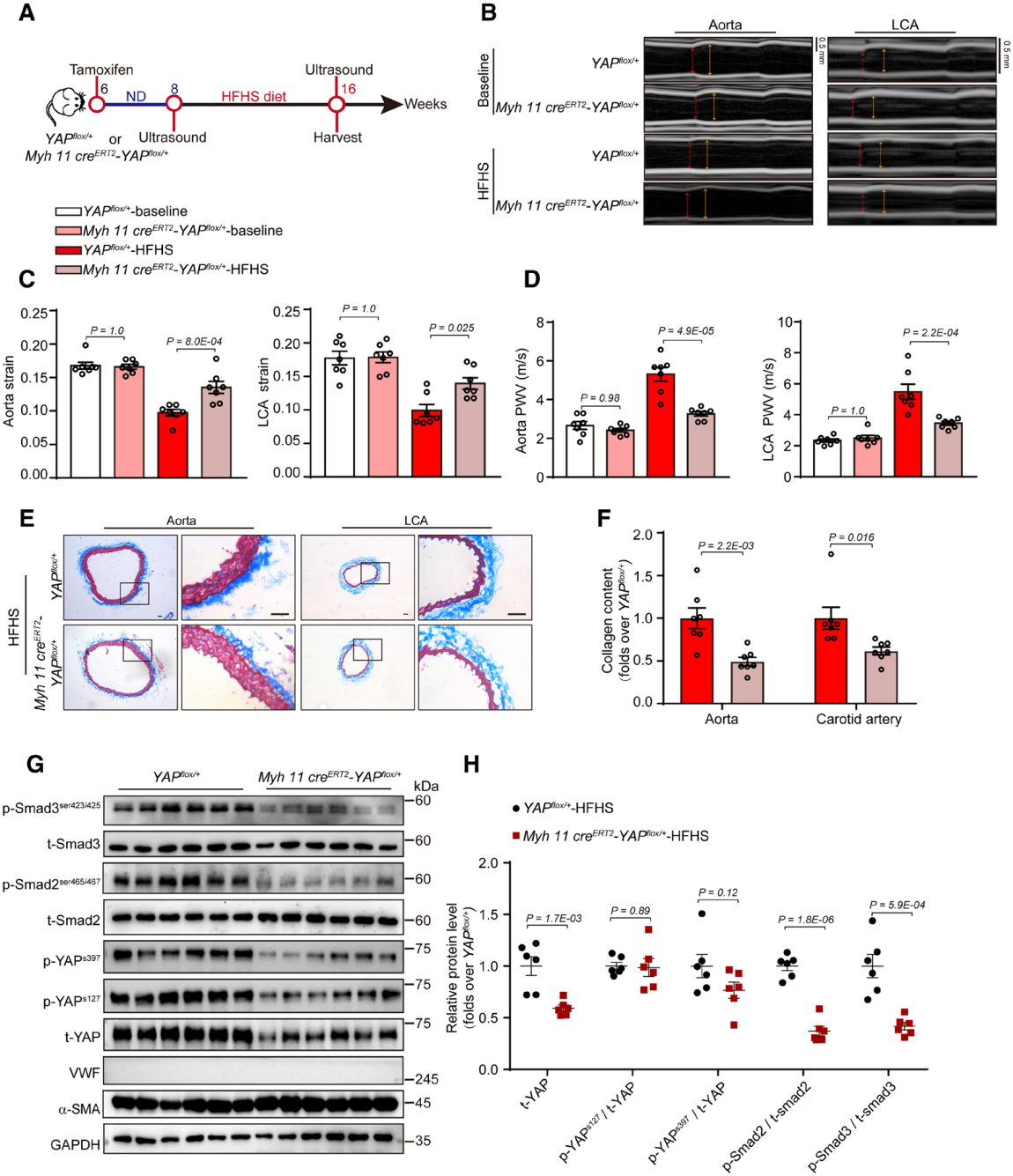
4. YAP蛋白在HFHS(高脂高糖)誘導的動脈硬化中有重要作用
研究者構建了平滑肌細胞特異性敲低YAP的小鼠模型,在高脂高糖飲食8周后,檢測血管硬化度。在喂食HFHS時,YAP敲低和未敲低組之間的附睪脂肪質量與體重的比率、血漿總膽固醇和甘油三酯水平、葡萄糖耐受和血壓均無顯著區別。YAP敲低組的動脈硬化不顯著,但有顯著更低的膠原蛋白聚集。此外,YAP敲低組的總YAP、磷酸化YAP、磷酸化Smad2和Smad3都較低。該結果表明,平滑肌細胞特異性敲低YAP能減輕高脂高糖(HFHS)誘導的動脈硬化。
5. YAP通過PPM1B依賴的機制在平滑肌細胞的TGFβ通路中發揮重要作用
為了研究YAP在TGFβ通路中的作用,研究者對人動脈平滑肌細胞(HASMC)進行了YAP的敲低和過表達。在TGFβ處理條件下,磷酸化Smad2/3在1h處達到峰值,隨后在2h開始降低,而YAP敲低組磷酸化Smad2/3都較對照組顯著低。在YAP過表達組,磷酸化Smad2/3都顯著升高。TGFβ誘導的Smad2的核定位以及ECM相關基因的mRNA表達在YAP敲低后均下調,而在YAP過表達后均上調。這些結果表明了一種YAP相關的激酶或磷酸酶調控了磷酸化Smad2/3的水平。
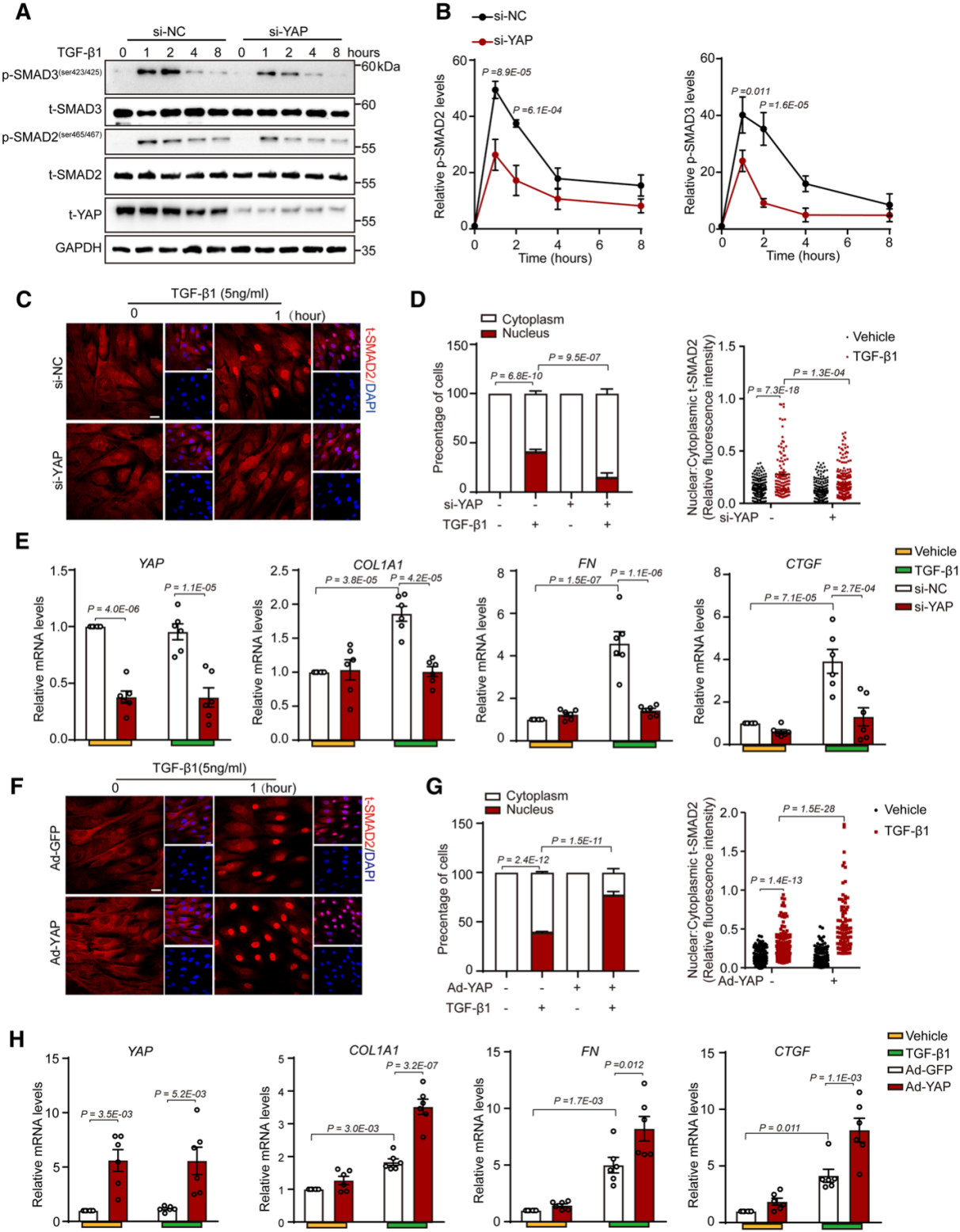
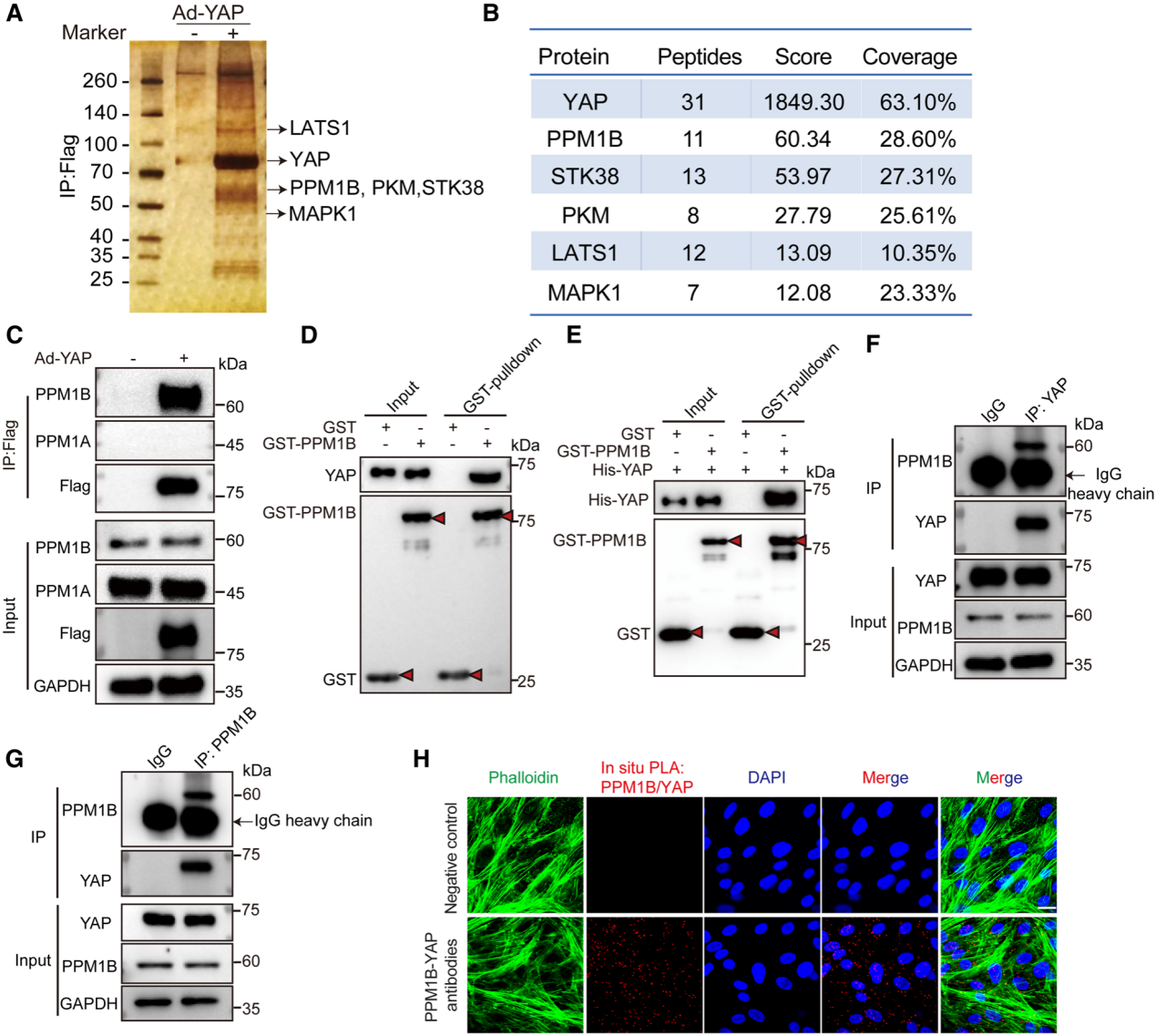
研究者通過進一步的實驗確認YAP過表達可以減緩Smad2/3的去磷酸化,并通過anti-FLAG-YAP CO-IP實驗和GST-PRM1B pull-down實驗確認了參與去磷酸化的磷酸酶為PPM1B。
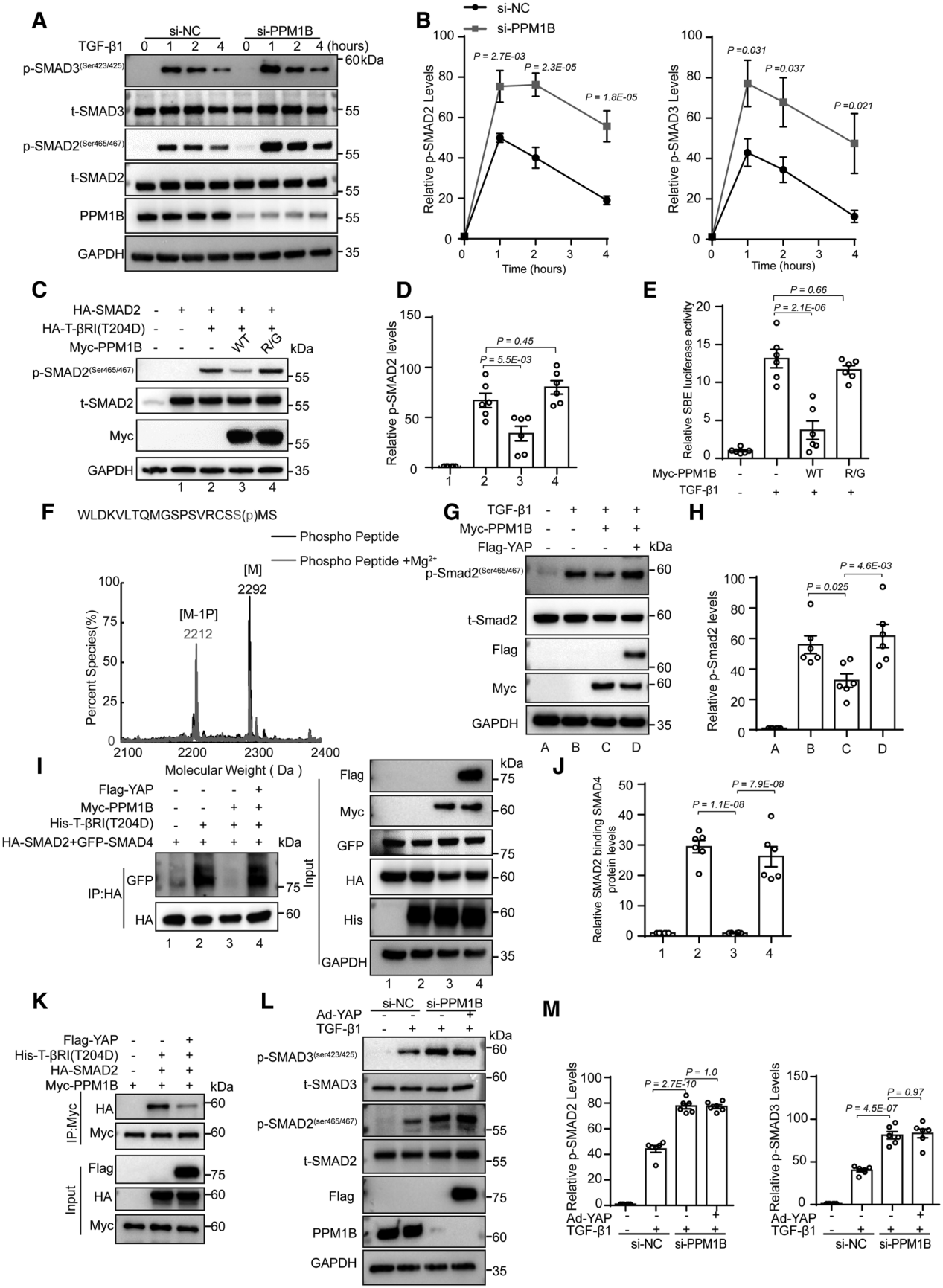
為了進一步探索PPM1B對Smad2/3的去磷酸化作用,研究者在HASMC(人動脈平滑肌細胞)中敲低或過表達PPM1B。發現在TGFβ處理的條件下,敲低PPM1B顯著上調磷酸化Smad2/3,而過表達PPM1B則顯著下調磷酸化Smad2/3。進一步,為了確認PPM1B對Smad2/3的去磷酸化是否是通過PPM1B的磷酸化活性來實現的,作者在HEK 293T細胞中表達TβR1 T204D(TGFβ受體1,204位蘇氨酸(T)突變為天冬氨酸(D),磷酸化激活型突變,該突變使TβR1 處于穩定的模擬磷酸化狀態,可持續磷酸化其底物Smad2/3,使之激活)。在表達TβR1 T204D的細胞中,野生型PPM1B可降低 Smad2/3的磷酸化,而磷酸酶活性失活型PPM1B(R179G,179位精氨酸(R)突變為甘氨酸(G))則不能降低Smad2/3的磷酸化。該結果表明,PPM1B對Smad2/3的去磷酸化依賴于PPM1B的磷酸酶活性。
接下來,研究者探究了YAP蛋白調控PPM1B依賴的Smad的去磷酸化。PPM1B過表達可以降低TGFβ1誘導的磷酸化Smad2的生成以及TβR1 T204D誘導的Smad2-Smad4復合體的形成。這兩種作用均可被YAP過表達逆轉。研究者同時發現,在PPM1B缺失的HASMC(人動脈平滑肌細胞)中,TGFβ1誘導的磷酸化Smad2/3的水平顯著升高,且YAP過表達無法進一步使Smad2/3磷酸化水平升高。該結果表明,YAP對磷酸化Smad2/3的作用依賴于PPM1B。
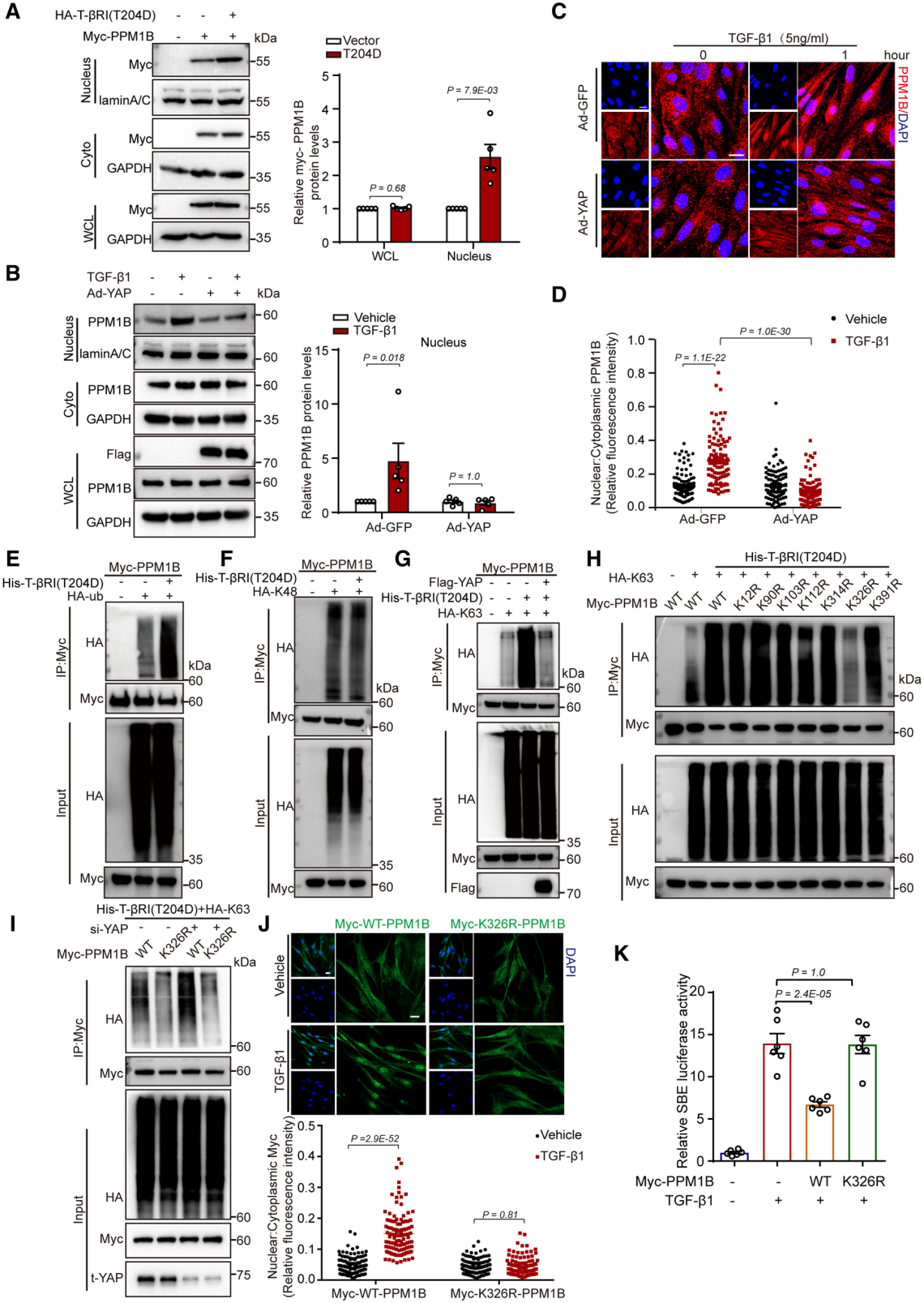
6. YAP抑制TGFβ誘導的PPM1B泛素化和核易位
內源性PPM1B主要位于細胞質中,而Smad2/3在細胞核中去磷酸化。因此,研究者假設在TGFβ通路激活條件下,PPM1B發生了核易位。最終的實驗結果也證明了研究者的假設:PPM1B在對TGFβ響應時易位到細胞核,執行了TGFβ誘導的磷酸化Smad2/3在細胞核中的去磷酸化,通過這樣的負反饋調節保證TGFβ的適當活性。此反饋調節由YAP控制。
翻譯后修飾對蛋白的穩定性、定位及構型有重要的影響。在PPM1B上僅報道過蛋白泛素化,因此研究者探索YAP調控PPM1B在細胞質中的停留是否是依賴于PPM1B的泛素化。最終的實驗表明,在YAP蛋白封閉了PPM1B K326(326位賴氨酸)的泛素化,隨后阻止了PPM1B易位到細胞核,最終減弱了對Smad2/3的去磷酸化。
7. YAP-CC結構域介PPM1B和YAP之間的物理相互作用
研究者進一步探索了YAP與PPM1B結合的結構域。通過表達不同截短的HA-YAP以及全長Myc-PPM1B,并進行pull-down實驗檢測YAP與PPM1B的結合情況。最終發現,當YAP缺失CC(coiled-coiled motif,卷曲螺旋基序)結構域時,anti-Myc免疫磁珠pull-down的產物中檢測不到YAP。隨后的實驗也表明,CC結構域對YAP與PPM1B的結合是必需的。緊接著,研究者將CC結構域分為coiled-coil區和非coiled-coil區,體外合成這兩條肽段并進行pull-down實驗,結果發現PPM1B選擇性與coiled-coil肽段結合。同樣地,研究者也明確了與YAP結合的區域是PPM1B的CD(C-terminal Domain)。研究者進一步將YAP CC結構域的關鍵氨基酸突變為甘氨酸(YAP-CC-G-mut),該突變廢除了YAP與PPM1B的結合,同時也無法增加TGFβ誘導下Smad2/3的磷酸化水平。此外,YAP-CC-G-mut也無法降低PPM1B K326的泛素化。以上結果表明,YAP-CC結構域和PPM1B CD對于TGFβ信號通路激活是非常重要的,因此也預示著它們是高脂高糖誘導的動脈硬化的潛在治療靶點。
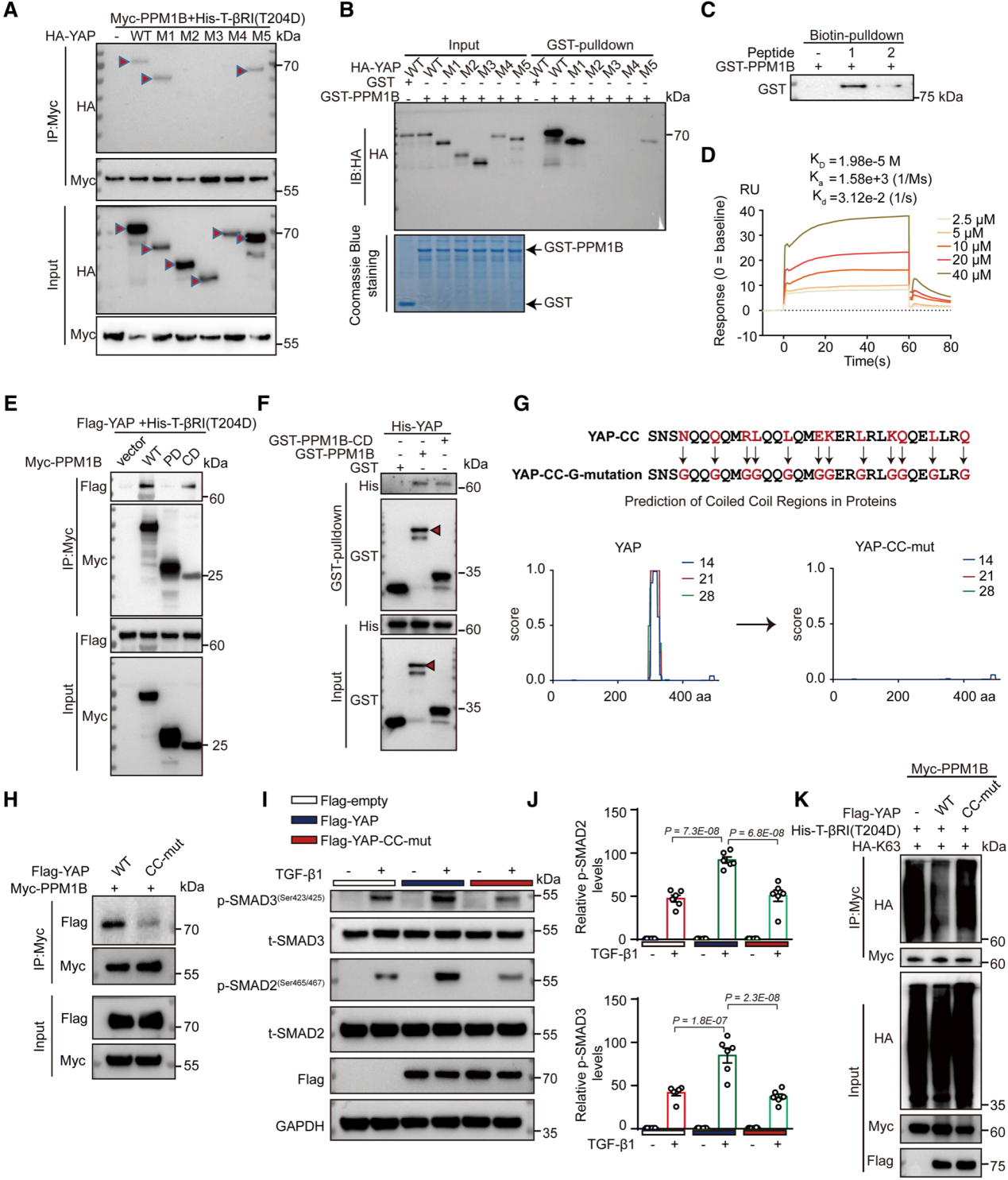
研究者在高脂高糖誘導的動脈硬化癥小鼠主動脈中膜中檢測到了TGFβ信號通路的激活,通過4D蛋白質組學檢測到了YAP蛋白的高表達。鑒于YAP蛋白與動脈硬化有關,研究者進一步探索了YAP在TGFβ信號通路中扮演的角色。最終的結果表明,YAP參與調控TGFβ/Smad2/3信號通路。機制上,YAP可以逆轉PPM1B泛素化,阻斷其轉位入核,最終使之不能將Smad2/3去磷酸化,從而保持了TGFβ信號通路的持續激活。結構上,YAP是通過其CC結構域與PPM1B的CD結合來發揮作用。作者的研究同時提示我們,靶向抑制YAP和PPM1B結合或可作為代謝綜合征患者預防動脈硬化的治療策略。
吉凱基因憑借多年在靶標篩選及驗證服務領域的技術積累,建立的標準化 、工程化 、系統化的GRP平臺,為中國研究型醫生提供科研服務,加快科研成果轉化。其中,多組學平臺包含蛋白質組學平臺和高通量測序平臺:
·蛋白質組學平臺擁有多臺timsTOF Pro、Exploris 480高精度質譜儀,專業的Spectronaut Plusar、Mascot等分析軟件,提供專業的4D、DIA、TMT、PRM、磷酸化修飾組、olink蛋白質組等檢測服務,強大的機器學習算法、IPA分析、蛋白基因組分析服務,系統的生物標志物、分子分型、藥物靶點、基因功能研究等解決方案,真正讓廣大研究型醫生的科研工作更省心、更省力、更高效;
·高通量測序平臺分為常規測序服務和單細胞測序服務:單細胞測序擁有10x和BD兩個平臺,提供單細胞RNA-seq、單細胞核測序、單細胞混樣RNA-seq、單細胞TCR/BCR、單細胞(RNA+ATAC)、空間轉錄組測序等服務;常規測序服務提供meRIP-seq(m6A/m1A/m7G/m5C 等RNA甲基化修飾測序)、acRIP-seq(ac4C RNA乙酰化修飾測序)、ATAC-seq、Ribo-seq(翻譯組測序) 、mRNA/miRNA/LncRNA/circRNA-seq、全轉錄組測序(兩文庫/三文庫)、外泌體miRNA/LncRNA-seq、WGS/WES、WGBS、RRBS、BSAS等服務。