α-突觸核蛋白與帕金森氏癥的病因與疾病發展有著密切的聯系。帕金森氏癥 (PD) 是一種神經衰退疾病,該疾病患者的大腦中可見α-突觸核蛋白聚集形成路易體。α-突觸核蛋白是由SNCA基因表達的一種分子量為 14-kDa 的蛋白。目前已知的α-突觸核蛋白的自然狀態可有很多種:可能是沒有折疊的單體1、折疊的四聚體、或者是處于和其他寡聚體動態共存的形式2。在帕金森疾病中,這些分子量小的α-突觸核蛋白會聚集形成原纖維、纖維以及路易體,從而導致神經元的病變和死亡。也有其他研究表明寡聚體和原纖維具有神經毒性3-6而路易小體可能有神經保護功能7。
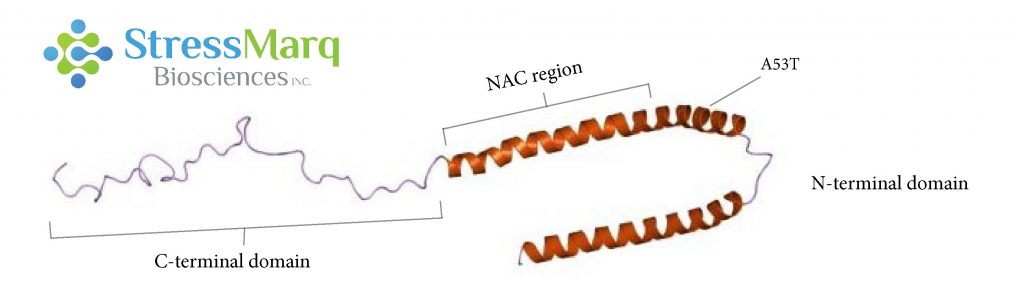
α-突觸核蛋白由三個主要區域:羧基末端區域、NAC區域、以及氨基末端區域。A53T 突變位于氨基末端區域。
A53T 是錯義點突變, 也就是導致了氨基酸的改變:第53 個氨基酸由丙氨酸突變為蘇氨酸。該突變是由于SNCA基因 209位置的鳥嘌呤變成了腺嘌呤 (G209A)8。A53T 突變與一種常染色體顯性遺傳的早發型 PD 相關,這種 PD 最先發現于意大利和希臘裔家族8,也見于一個韓國裔家族9。 A53T 變異導致該疾病的發病年齡比較早8。雖然大部分的 PD 病例是散發型的,并非遺傳病,而且也不涉及 A53T 突變, 但是研究 A53T 突變可以幫助科研學者們更好的了解α-突觸核蛋白的聚集和發展機制,從而研發出更好的疾病模型和治療方案。
?
?
?
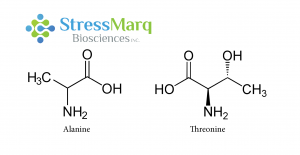
丙氨酸和蘇氨酸有相似的結構,但是α-突觸核蛋白中的丙氨酸替換成蘇氨酸對α突觸核蛋白的纖維原聚集有很大的作用。
A53T 變異只涉及到單一氨基酸改變,而且丙氨酸和蘇氨酸結構上非常相近,那么為什么A53T突變型有這么強的聚集效果呢?這是因為A53T 和其他致病突變都發生在α-突觸核蛋白的氨基末端,理論模型顯示 A53T 突變會使α-突觸核蛋白的 NAC 區域和氨基或羧基區域的長程相互作用消失,導致 beta折疊的加速形成10。NMR 測量數據顯示 A53T 突變可以延長并穩定在寡聚化和纖維化中起著重要作用的 beta折疊結構11。因此A53T 突變型能夠更快聚集alpha突觸核蛋白,有更明顯的致病效果。下面的硫黃素T檢測曲線可以明顯看出A53T突變型的聚集效果明顯強于非突變型α-突觸核蛋白。
A53T 變異只涉及到單一氨基酸改變,而且丙氨酸和蘇氨酸結構上非常相近,那么為什么A53T突變型有這么強的聚集效果呢?這是因為A53T 和其他致病突變都發生在α-突觸核蛋白的氨基末端,理論模型顯示 A53T 突變會使α-突觸核蛋白的 NAC 區域和氨基或羧基區域的長程相互作用消失,導致 beta折疊的加速形成10。NMR 測量數據顯示 A53T 突變可以延長并穩定在寡聚化和纖維化中起著重要作用的 beta折疊結構11。因此A53T 突變型能夠更快聚集alpha突觸核蛋白,有更明顯的致病效果。下面的硫黃素T檢測曲線可以明顯看出A53T突變型的聚集效果明顯強于非突變型α-突觸核蛋白。
?
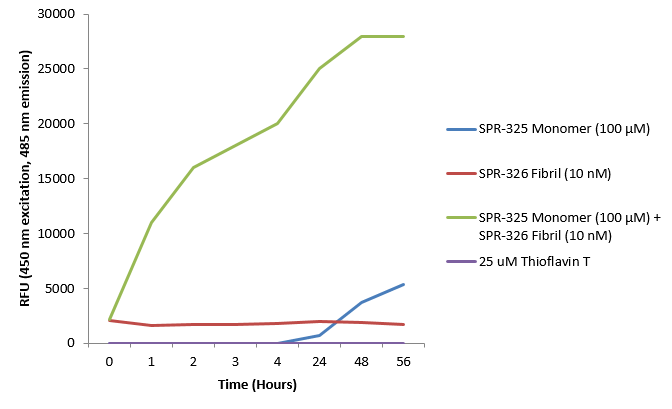 |
硫黃素 T 是一種熒光染料, 可以綁定富含beta折疊的結構, 例如 α-Syn 聚合體. 綁定之后, 硫黃素T光譜會發生紅移, 熒光強度會增強. A53T α-Syn蛋白單體 (SPR-325)的硫黃素 T 發射光曲線顯示了有限的熒光強度增強 (相關于α-Syn 蛋白聚合體). 而 10 nM 活性 α-Syn 聚合體 (SPR-326) 與100 μm 活性 α-Syn 單體 (SPR-325) 混合時熒光強度明顯增強, 這是由于聚合體催化了活性單體形成更多聚集體PFFs. Thioflavin T λex = 450 nm, λem = 485 nm. |
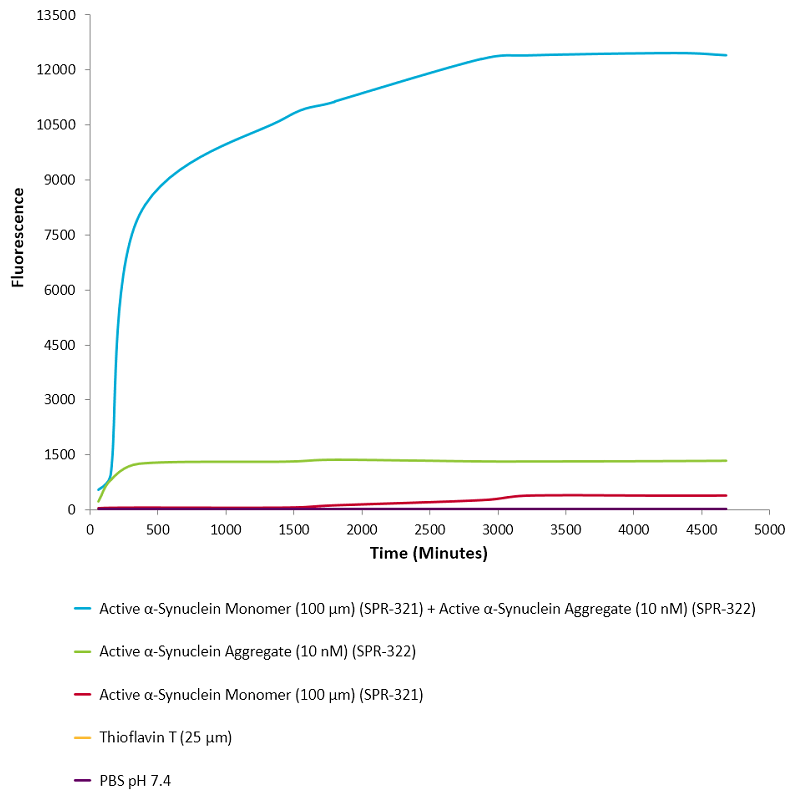 |
活性突觸核蛋白 (α-Syn) 聚合體 (SPR-322) 催化活性單體(SPR-321)形成新的聚合體. 硫黃素 T 是一種熒光染料, 可以綁定富含beta折疊的結構, 例如 α-Syn 聚合體. 綁定之后, 硫黃素T光譜會發生紅移, 熒光強度會增強. 左側硫黃素 T 發射光曲線展示了四種試驗對象隨時間增強的熒光強度(相關于α-Syn 蛋白聚合體),其中 10 nM 活性 α-Syn 聚合體(SPR-322) 與100 μm 活性 α-Syn 單體 (SPR-321) 混合物熒光增強最明顯, 另外幾組對比分別是活性 α-Syn 聚合物 (SPR-322) 、活性 α-Syn 單體 (SPR-321) 以及 Thioflavin T λex = 450 nm, λem = 485 nm. |
A35T突變型 alpha 突觸核蛋白新品上市半價促銷,活動時間2019年3月1日至4月30日,折扣碼 A35T50.
| 活性重組人 A53T 突變型 Alpha Synuclein 蛋白單體 (1 型) | SPR-325B | 100 μg |
| SPR-325C | 2 x 100 μg | |
| SPR-325E | 5 x 100 μg | |
| 活性重組人 A53T 突變型 Alpha Synuclein 蛋白 PFFs (1 型) | SPR-326B | 100 μg |
| SPR-326C | 2 x 100 μg | |
| SPR-326E | 5 x 100 μg |
參考文獻:
- Fauvet B, et al. alpha-Synuclein in central nervous system and from erythrocytes, mammalian cells, and Escherichia coli exists predominantly as disordered monomer. J Biol Chem. 2012;287:15345–64.
- Dehay B, Bourdenx M, Gorry P, et al. Targeting α-synuclein for treatment of Parkinson’s disease: mechanistic and therapeutic considerations. Lancet Neurol. 2015;14(8):855-866.
- Karpinar DP, et al. Pre-fibrillar alpha-synuclein variants with impaired beta-structure increase neurotoxicity in Parkinson’s disease models. EMBO J. 2009;28:3256–68.
- Winner B, et al. In vivo demonstration that alpha-synuclein oligomers are toxic. Proc Natl Acad Sci U S A. 2011;108:4194–9.
- Cremades N, et al. Direct observation of the interconversion of normal and toxic forms of alpha-synuclein. 2012;149:1048–59.
- Danzer KM, et al. Different species of alpha-synuclein oligomers induce calcium influx and seeding. J Neurosci. 2007;27:9220–32.
- Tanaka M, et al. Aggresomes formed by alpha-synuclein and synphilin-1 are cytoprotective. J Biol Chem. 2004;279:4625–31.
- Polymeropoulos, M. H. Mutation in the -Synuclein Gene Identified in Families with Parkinson’s Disease. Science, 1998;276(5321), 2045–2047. doi:10.1126/science.276.5321.2045
- Ki C.S. Stavrou E.F. Davanos N. Lee W.Y. Chung E.J. Kim J.Y. Athanassiadou A. The Ala53Thr mutation in the alpha-synuclein gene in a Korean family with Parkinson disease. Clin Genet. 2007 May;71(5):471-3.
- Coskuner, O., Wise-Scira, O. Structures and Free Energy Landscapes of the A53T Mutant-Type α?Synuclein Protein and Impact of A53T Mutation on the Structures of the Wild-Type α?Synuclein Protein with Dynamics. ACS Chem. Neurosci. 2013, 4, 1101?
- Russel, R., Eliezer, D. Residual structure and dynamics in Parkinson’s disease-associated mutants of alpha-synuclein. J Biol Chem. 2001 Dec 7;276(49):45996-6003.