nature子刊|北大張迪教授團隊:賴氨酸L-乳酸化是糖酵解誘導的主要乳酸化異構體

英文標題:Lysine L-lactylation is the dominant lactylation isomer induced by glycolysis
中文標題:賴氨酸L-乳酸化修飾(lysine L-lactylation)是糖酵解誘導產生的主要乳酸化異構體
發表期刊:nature chemical biology
影響因子:13.7
文章簡介
芝加哥大學趙英明教授、威斯康星大學麥迪遜分校的李靈軍教授、四川大學的戴倫治教授以及北京大學的張迪教授等團隊在國際權威期刊nature chemical biology上發表了題為“Lysine L-lactylation is the dominant lactylation isomer induced by glycolysis”的最新研究成果。該研究開發了區分賴氨酸乳酸化三種異構體的技術,證實賴氨酸L-乳酸化(KL-la)是組蛋白上的主要異構體,且是糖酵解和瓦伯格效應的主要響應者,同時明確其前體乳酸輔酶 A(lactyl-CoA)與KL-la水平正相關。
研究背景
賴氨酸乳酸化修飾是由L-乳酸驅動的新型蛋白質翻譯后修飾(PTM),在代謝與表觀遺傳調控的關聯中具有關鍵意義,但其存在的三種異構體——賴氨酸L-乳酸化(KL-la)、d-乳酸基-賴氨酸(KD-la)與N-ε-(羧乙基)-賴氨酸(Kce),因分子量相同、結構相似,常規 HPLC-MS技術難以區分,導致三者在瓦伯格效應及核內修飾功能研究中長期混淆,嚴重阻礙對其真實生物學功能的認知。
前期研究已明確:KL-la由糖酵解衍生的L-乳酸驅動形成,推測乳酸輔酶A(lactyl-CoA)是其形成的高能中間物,p300可催化組蛋白KL-la形成,HDAC1-3、SIRT2參與去修飾;KD-la由蛋白質與乙二醛酶途徑產物LGSH非酶促生成;Kce是糖酵解副產物MGO與賴氨酸的加合物,且組蛋白上Kce水平遠低于精氨酸修飾。但關鍵問題仍待解決:三種異構體在組蛋白上的主要存在形式不明確;三者對糖酵解動態變化的響應模式未系統探究;lactyl-CoA在細胞內的存在、代謝來源及與KL-la的關聯性缺乏直接證據,且學界對三者生化機制與功能差異存在爭議,部分研究質疑KL-la真實性及lactyl-CoA存在。因此,開發可靠的異構體區分方法、厘清其與糖酵解的關系,成為深入理解代謝-表觀調控軸的迫切需求。
技術路線
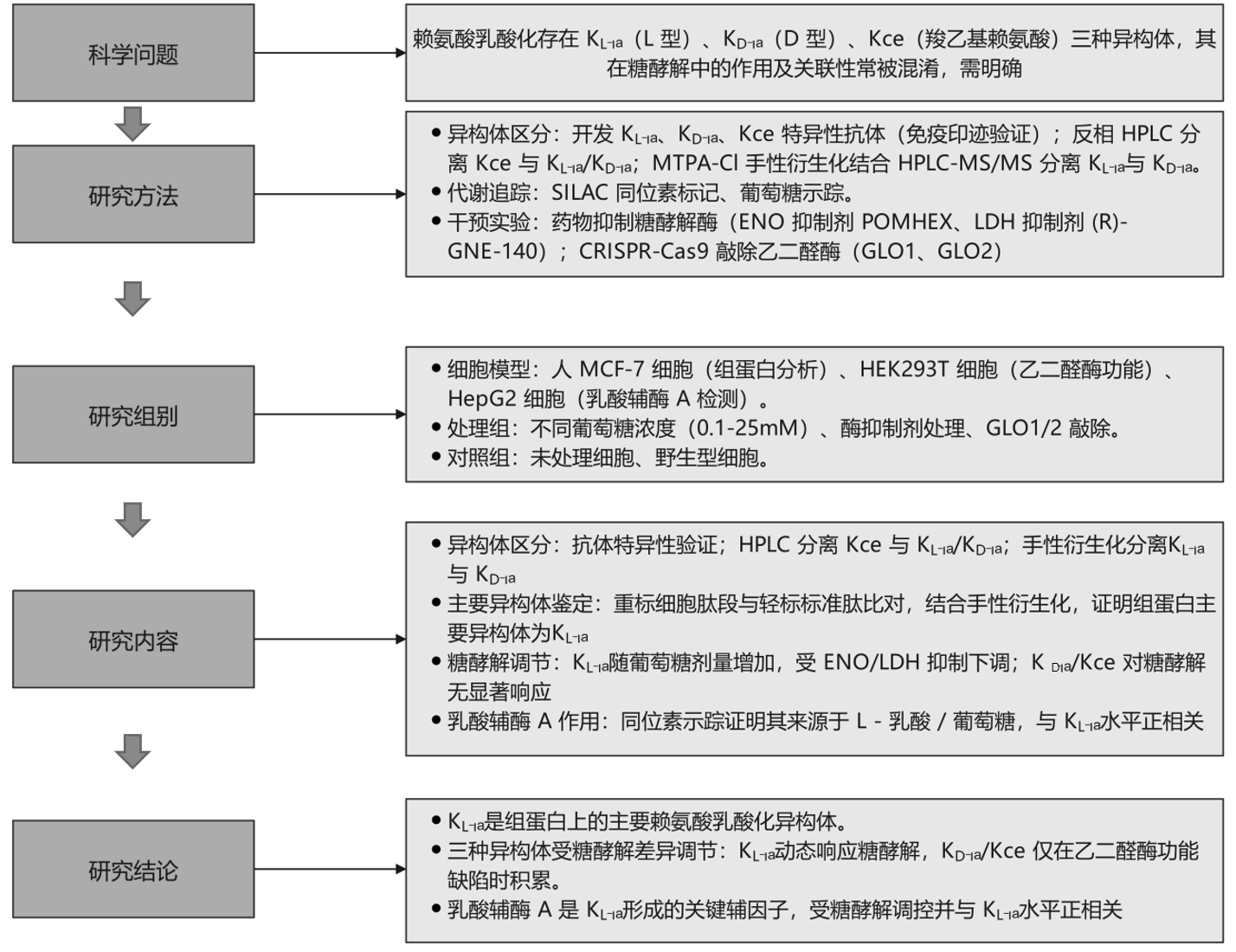
研究結果
1、高效液相色譜(HPLC)可將Kce肽與KL-la或KD-la肽分離
翻譯后修飾(PTM)特異性抗體可區分賴氨酸L-乳酸化(KL-la)、賴氨酸D-乳酸化(KD-la)和N-ε-(羧乙基)-賴氨酸(Kce)(圖1)。本研究中,研究人員開發了第二代兔單克隆泛抗KL-la抗體,以及泛抗KD-la和泛抗Kce抗體。為探究這些試劑的有效性,研究人員進行了免疫印跡實驗(圖2a),檢測合成肽庫(圖2b-d)、化學修飾的牛血清白蛋白(BSA)(圖2e-g)和合成的序列特異性組蛋白肽(圖2h-j)。結果顯示,泛抗KL-la、泛抗KD-la和泛抗Kce抗體對各自對應的PTM具有特異性,比對其他修飾的偏好性高至少50倍(圖2b-j)。
隨后該研究利用HPLC實驗區分KL-la、KD-la和Kce,結果發現盡管帶有KL-la或KD-la的肽對無法分離,但通過反相HPLC可以輕松區分它們與帶有Kce的對應肽(圖3a-b)。
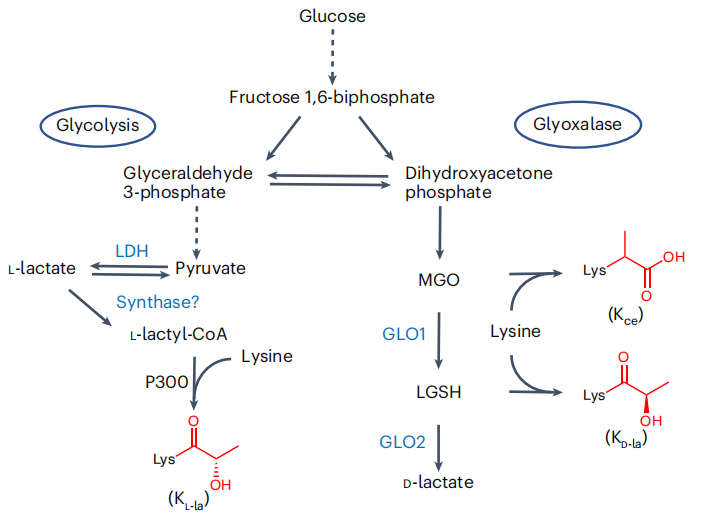
圖1. KL-la、KD-la和Kce是與糖酵解相關的結構異構體
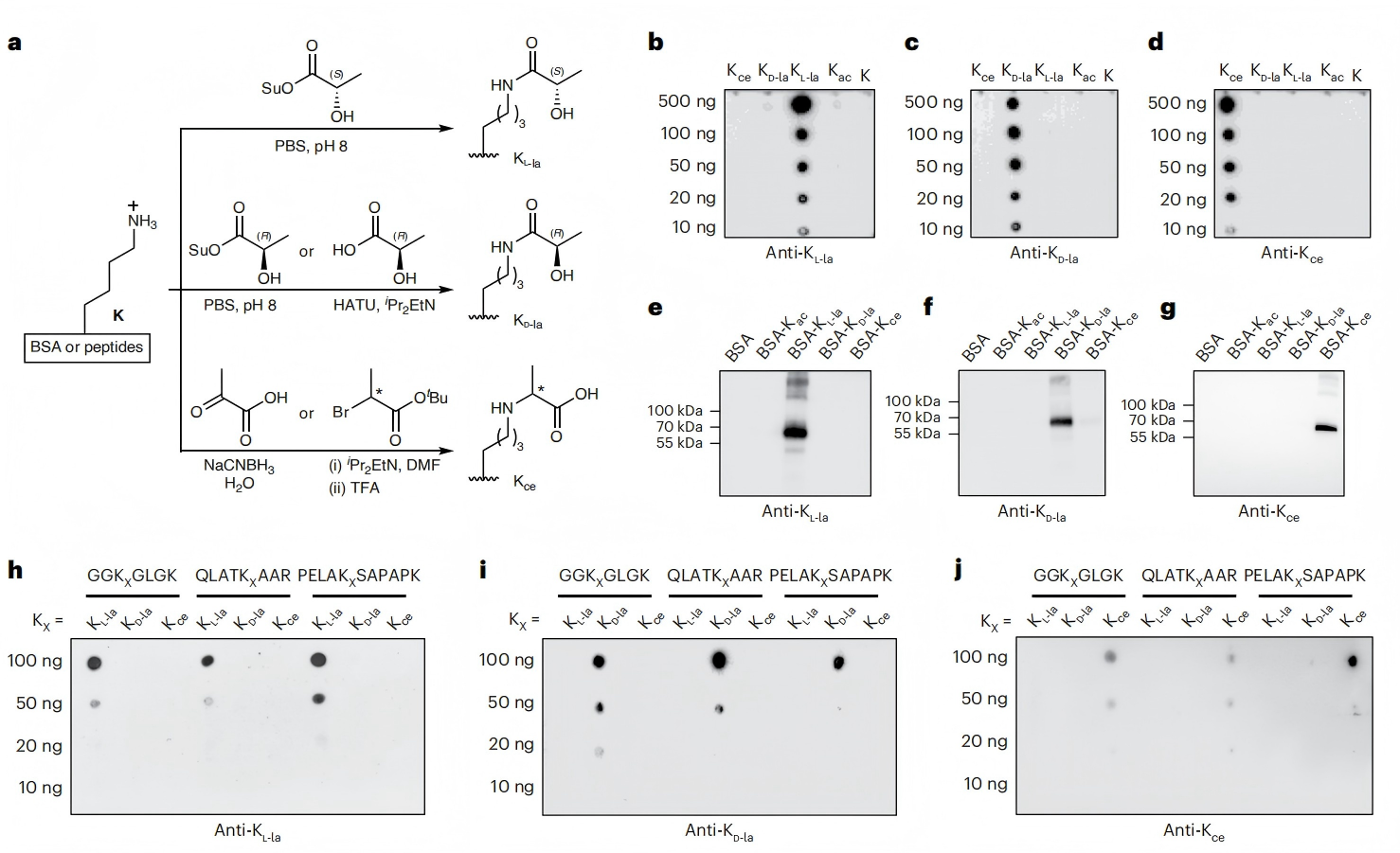
圖2. PTM特異性抗體可區分KL-la、KD-la和Kce
2、手性反應結合HPLC實現KL-la與KD-la的分離
研究人員利用經典的Mosher酸氯化物(MTPA-Cl)手性衍生化反應,該反應可以放大手性差異,從而提高色譜分離的可能性(圖3c)。為探究這種可能性,研究人員首先將肽消化成氨基酸,再經過MTPA-Cl衍生化后,進行HPLC-MS/MS分析(圖3d)。結果發現,(MTPA)2-KL-la和(MTPA)2-KD-la能得到很好的分離(圖3e),且在MS/MS中具有相同的碎片模式(圖3f)。
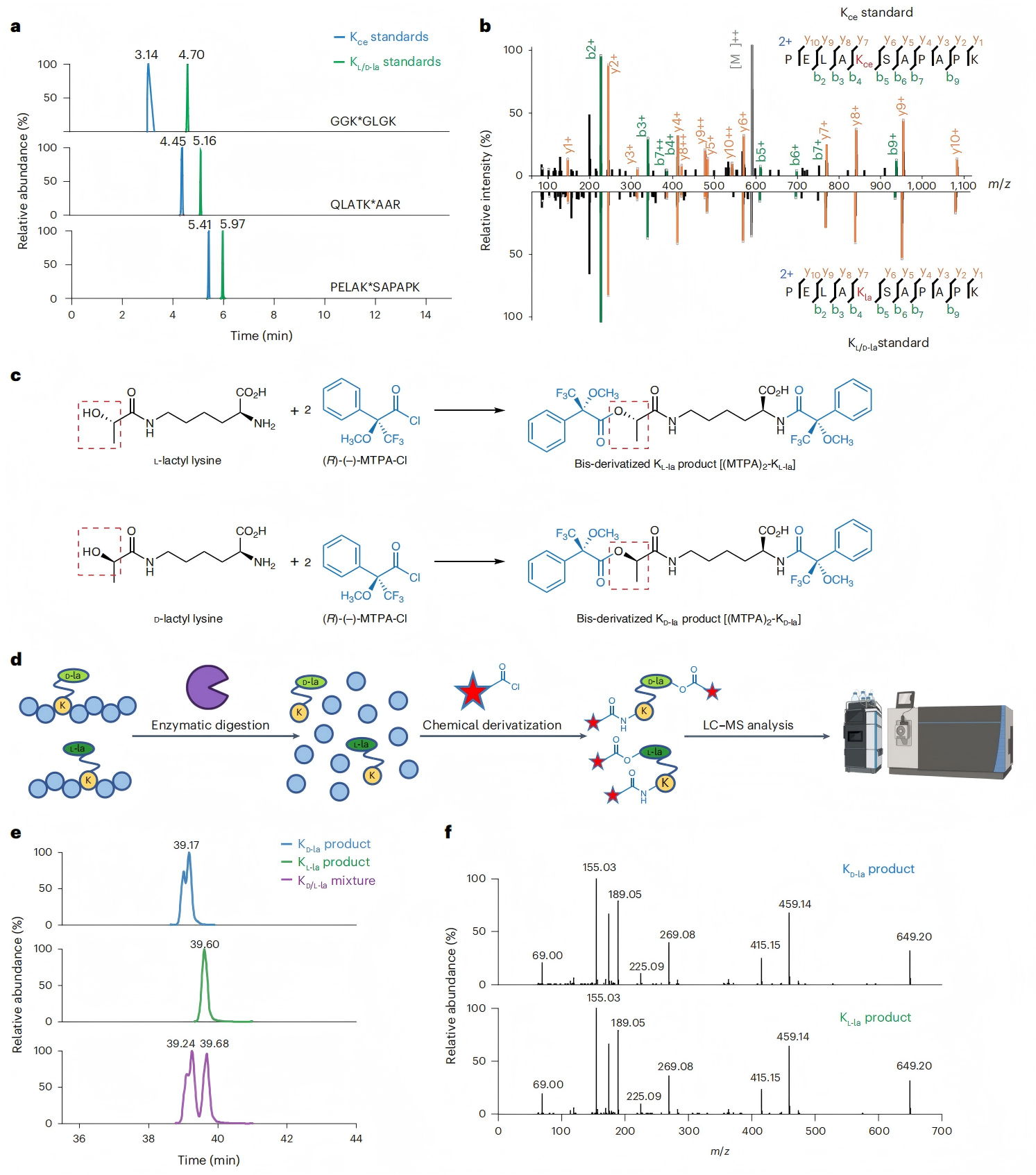
圖3. 通過HPLC-MS/MS分離含KL-la、KD-la和Kce的多肽
3、KL-la是組蛋白上的主要乳酸化異構體
研究證實,KL-la是組蛋白上的主要修飾,而非KD-la或Kce。 研究人員通過“重標”培養基(含K8[¹³C?,¹?N?]標記L-賴氨酸)培養人MCF-7細胞,富集組蛋白修飾肽段后,與“輕標”(K0[¹²C?,¹?N?])KL-la、KD-la和Kce標準肽混合,進行HPLC-MS/MS 分析(圖4a)。結果顯示,細胞來源肽段與KL/D-la標準肽保留時間及碎片模式一致,與Kce標準肽不一致,表明組蛋白主要修飾為KL/D-la而非Kce(圖4b)。
為明確是KL-la還是KD-la,研究人員將富集肽段消化為氨基酸后進HPLC-MS/MS分析,結果顯示,細胞來源產物與KL-la產物共洗脫,證明組蛋白乳酸化主要為L型(圖4c-d)。 基于SILAC的定量質譜驗證顯示,高葡萄糖誘導62個KL-la修飾組蛋白肽段,核心組蛋白KL-la對高葡萄糖響應更顯著,且未檢測到KD-la或Kce修飾(圖4e-f)。 綜上數據,細胞組蛋白的三種賴氨酸乳酸化結構異構體中,KL-la是主要異構體。
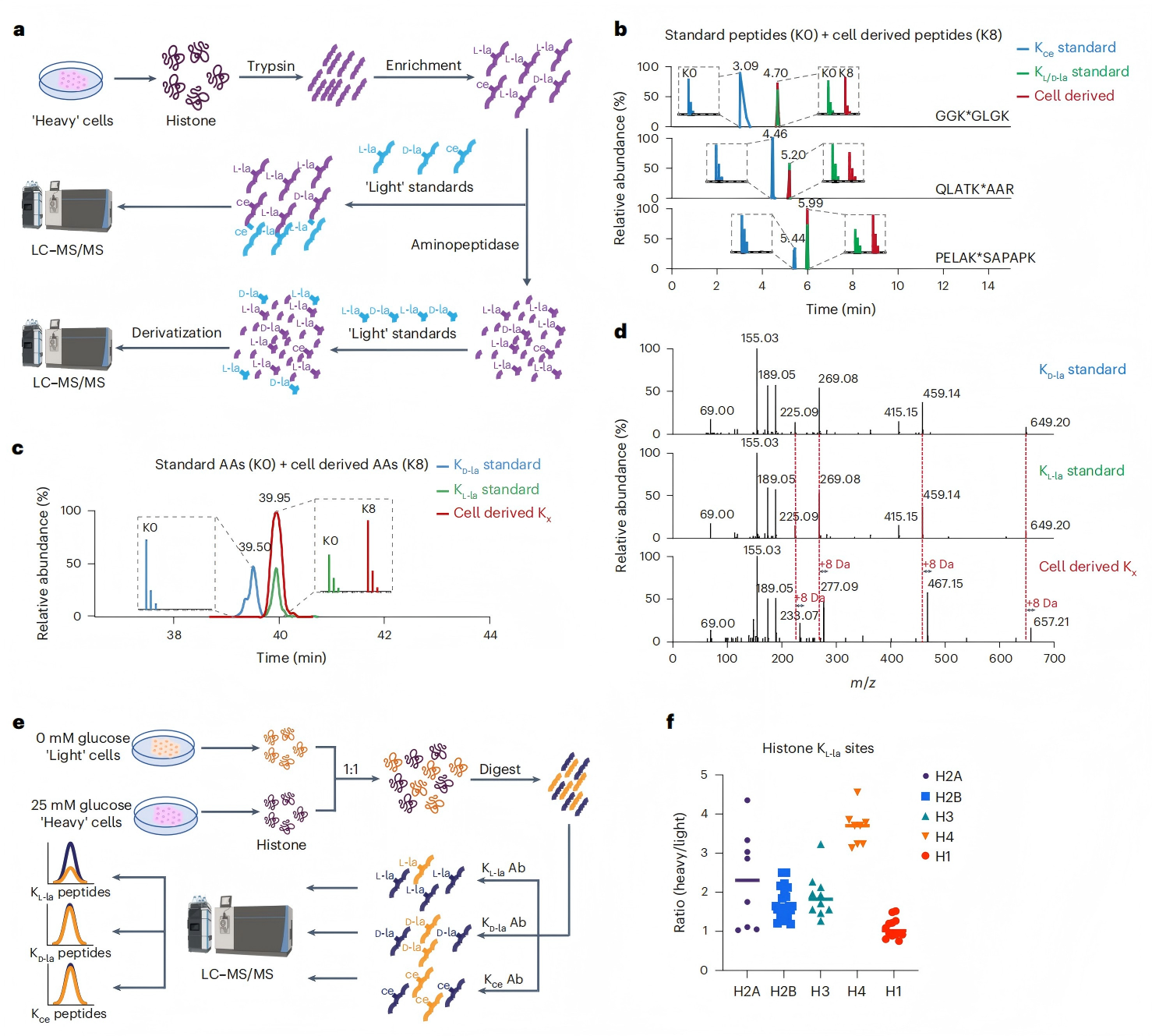
圖4. KL-la是細胞組蛋白上最常見的翻譯后修飾
4、糖酵解對KL-la、KD-la和Kce的調節存在差異
KL-la、KD-la和Kce均由糖酵解中間產物作為前體形成(圖5a)。研究人員在含不同葡萄糖濃度的培養基中培養人MCF-7細胞,通過蛋白質印跡法檢測發現,組蛋白和非組蛋白上的整體KL-la水平被葡萄糖以劑量依賴方式誘導,而KD-la和Kce水平對細胞外葡萄糖濃度基本無響應(圖5b)。
為探究糖酵解關鍵步驟對三種異構體的調控作用,研究人員通過抑制烯醇化酶(ENO)或乳酸脫氫酶(LDH)干擾糖酵解過程(圖5a),結果顯示,ENO抑制劑POMHEX以劑量依賴方式降低整體KL-la水平,同時使KD-la水平增加,Kce水平受輕微影響(圖5c);而LDH抑制劑(R)-GNE-140僅特異性降低整體KL-la水平,對KD-la和Kce無影響(圖5d)。 為進一步明確乙二醛酶途徑的作用,研究人員將GLO1缺陷、GLO2缺陷和GLO1/2雙缺陷的人胚胎腎(HEK)293T細胞,分別培養于含不同葡萄糖濃度的培養基中。檢測結果表明,整體KL-la對葡萄糖濃度有靈敏響應,但不受乙二醛酶(GLO1/GLO2)調節;KD-la僅在GLO2缺失時被誘導,對葡萄糖濃度變化敏感性低;僅GLO1缺失時Kce水平未增加(圖5e)。 綜上,KL-la、KD-la和Kce在糖酵解過程中受不同調節,乙二醛酶途徑可防止細胞中KD-la和Kce的形成,而不影響對糖酵解敏感的KL-la。
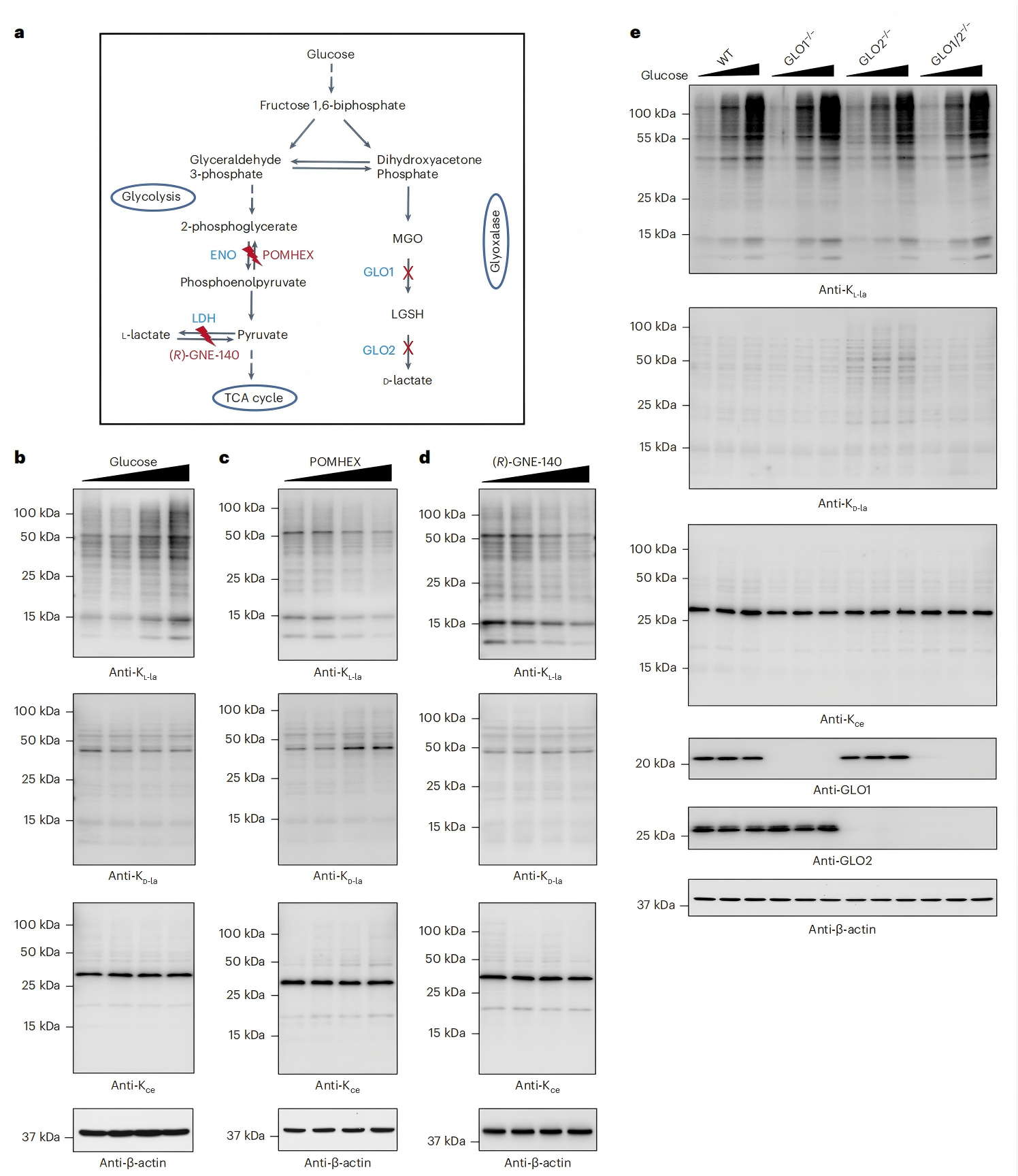
圖5. 糖酵解對KL-la、KD-la和Kce的差異調控
5、乳酸輔酶A與KL-la在響應糖酵解時相關
研究人員提出乳酸輔酶A是KL-la形成的輔因子,且乙酰轉移酶p300可利用化學合成的乳酸輔酶A作為體外酶促組蛋白KL-la反應的輔因子。為驗證該結論,實驗首先確認了培養細胞中乳酸輔酶A的存在(圖6a-b)。通過¹³C?-L-乳酸和U-¹³C?-D-葡萄糖同位素示蹤實驗發現,乳酸輔酶A和乙酰輔酶A均能被外源性¹³C?-L-乳酸快速標記,4小時后標記減少,且乳酸輔酶A的最大標記遠高于乙酰輔酶A,丙酰輔酶A未被標記(圖6c);用U-¹³C?-D-葡萄糖處理細胞,結果顯示,乙酰輔酶A標記比乳酸輔酶A快得多,丙酰輔酶A未被標記(圖6d),表明乳酸輔酶A可從L-乳酸和葡萄糖代謝衍生而來。 在ENO或LDH被阻斷的細胞中測量顯示,POMHEX處理后乳酸輔酶A水平顯著降低;LDH缺陷細胞或經(R)-GNE-140處理的細胞中,乳酸輔酶A水平也遠低于對照細胞(圖6e-f),表明乳酸輔酶A受糖酵解動態調節,并與KL-la水平呈正相關。
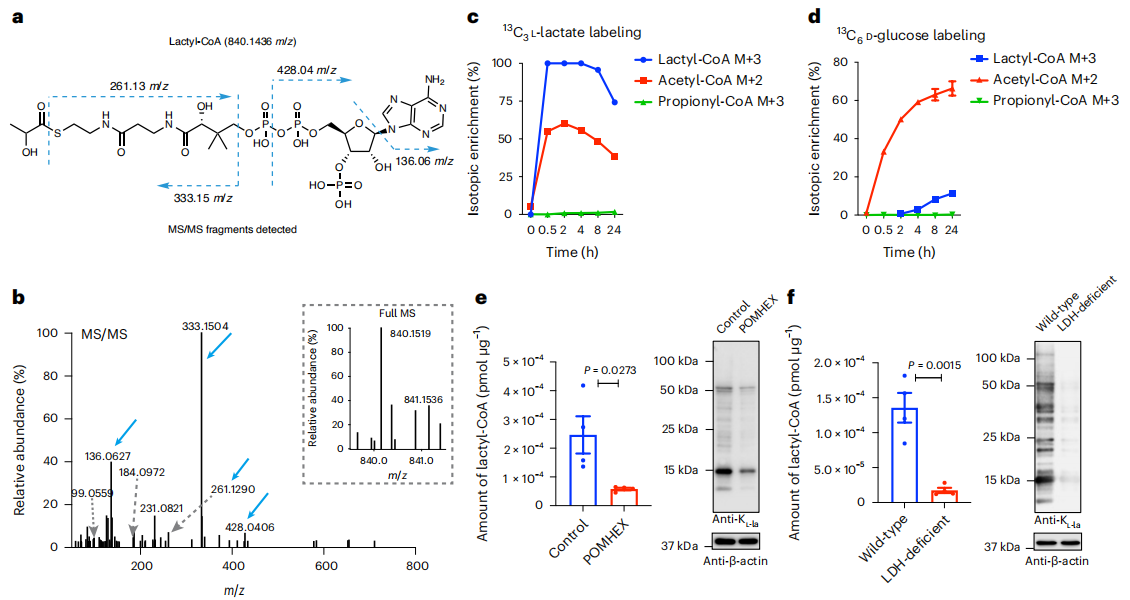
圖6. 在糖酵解過程中,乳酰輔酶A與KL-la呈正相關
研究結論
本研究為深入理解蛋白質翻譯后修飾的復雜性奠定了基礎,特別是在糖酵解相關的代謝調控和表觀遺傳調控交叉領域。通過明確KL-la作為主要響應糖酵解的異構體,為后續研究其在生理和病理過程中的功能作用提供了關鍵依據,同時也為開發針對特定異構體的調控手段開辟了新的方向。
乳酸化修飾(Lactylation):是一種蛋白質翻譯后修飾,乳酸(Lactic)和輔酶A(CoA)在特定酶的作用下結合,形成乳酰輔酶A(Lactyl-CoA),作為乳酸化修飾的供體,再由特定的乳酸轉移酶將乳酸基團轉移到蛋白質的賴氨酸殘基上,發生乳酸化修飾。這一過程改變了蛋白質的電荷狀態、結構和功能,從而影響蛋白質的活性、定位以及與其他分子的相互作用,進一步調控廣泛的細胞生理過程和病理狀態。
百趣生物提供“乳酸化修飾”等專業技術服務,助您快速解鎖研究瓶頸。
?
END
快樂小包子 撰文
Tang 校稿