本期將帶您深入基因編輯正史的核心:一把源自細菌與病毒30億年攻防戰的"分子剪刀"——CRISPR/Cas9技術,它正以納米級精度重寫生命密碼。這場顛覆生物醫學領域的革命,其開端竟隱藏在一次被忽視的實驗室意外中!
從“垃圾序列”到基因魔剪的蛻變
1987年,日本科學家石野良純在解析大腸桿菌基因組中某些酶功能時,意外地在某個片段中發現了一些神秘的重復序列(即后來的CRISPR序列),但他并未深究這些看似無規律但又極具規律的片段,僅留下“值得深入探討”幾個字,因此這些奇特的序列在隨后的十八年里一直沉睡于科學文獻中。

神秘的重復序列(來自 Ishino, Y , et al. 1987)
此后,西班牙和荷蘭等各地科學家也陸續在細菌和古細菌中發現新的重復序列,并發現這些重復序列和間隔序列的排列方式、重復序列重復出現的次數都可以表現出很大差異。因此Ruud Jansen在2002年將具有以上特征的DNA片段命名為CRISPR(Clustered Regularly Interspaced Short Palindromic Repeats,成簇規律性間隔短回文重復序列),但依舊無法解釋它的作用。
直到2005年,西班牙微生物學家Francisco Mojica發表的文章才揭開了這些序列的真實身份:他證實CRISPR間隔序列匹配噬菌體DNA,提出其作為細菌“免疫記憶庫”的假說。細菌通過將病毒DNA片段嵌入自身基因組,形成“分子通緝令”,當相同病毒再次來襲時,便能精準識別并摧毀入侵者。這一發現開啟了基因編輯的新紀元!
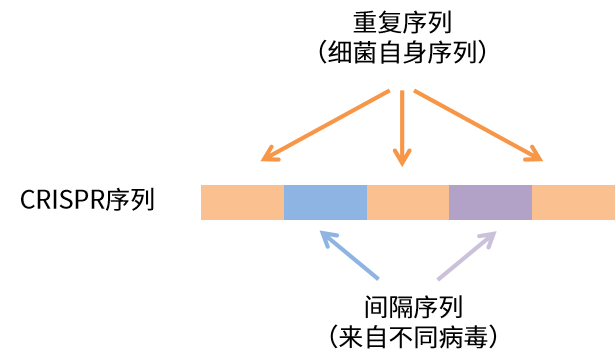
2011年,Emmanuelle Charpentier團隊發現CRISPR/Cas9系統的核心組分:crRNA(由CRISPR轉錄而來)與tracrRNA(反式激活crRNA)結合,指導Cas9蛋白切割目標DNA。
CRISPR/Cas9系統(來自Emmanuelle Charpentier,2012)
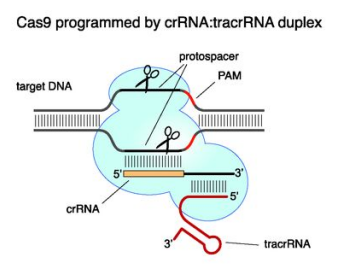
2012年,Jennifer Doudna與Emmanuelle Charpentier在《Science》發表載入基因編輯史冊的里程碑研究:她們將復雜的CRISPR系統簡化為Cas9蛋白+sgRNA 的二元組合,開發了第三代基因編輯工具——CRISPR/Cas9技術。只需修改sgRNA的20個堿基序列,就能引導Cas9精準切割任意DNA靶點。更因此獲得了2020年的諾貝爾化學獎!

(來自諾貝爾獎官網)
次年,張鋒發表了利用CRISPR技術實現哺乳動物細胞基因編輯的文章,標志著CRISPR技術從理論到實踐的一大飛躍。后續科學家們的不斷深入探索研究,CRISPR技術逐漸完善,正朝著基因編輯的時代邁進。
細菌免疫系統如何變身基因魔剪?
CRISPR系統的本質是RNA引導的DNA切割引擎,其納米級精準性源于三大核心組件的精密協作:
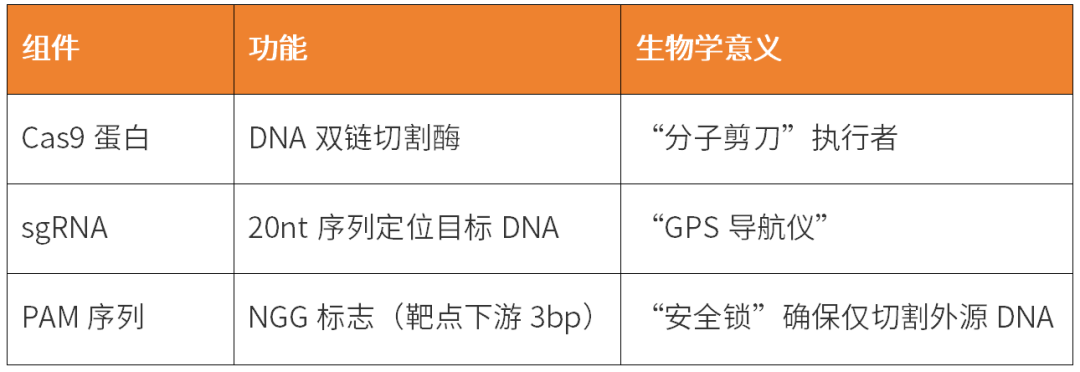
當這些組件協同工作時,將啟動四步精密機制:
1.組裝:Cas9與sgRNA結合形成核糖核蛋白復合體(RNP);
2.掃描:sgRNA通過堿基互補配對在基因組中搜尋目標;
3.解鎖:Cas9識別PAM序列(NGG)后解旋DNA雙鏈;
4.切割:Cas9蛋白的HNH結構域切斷互補鏈,RuvC結構域切斷非互補鏈。獲得斷裂的雙鏈DNA(DSB)。
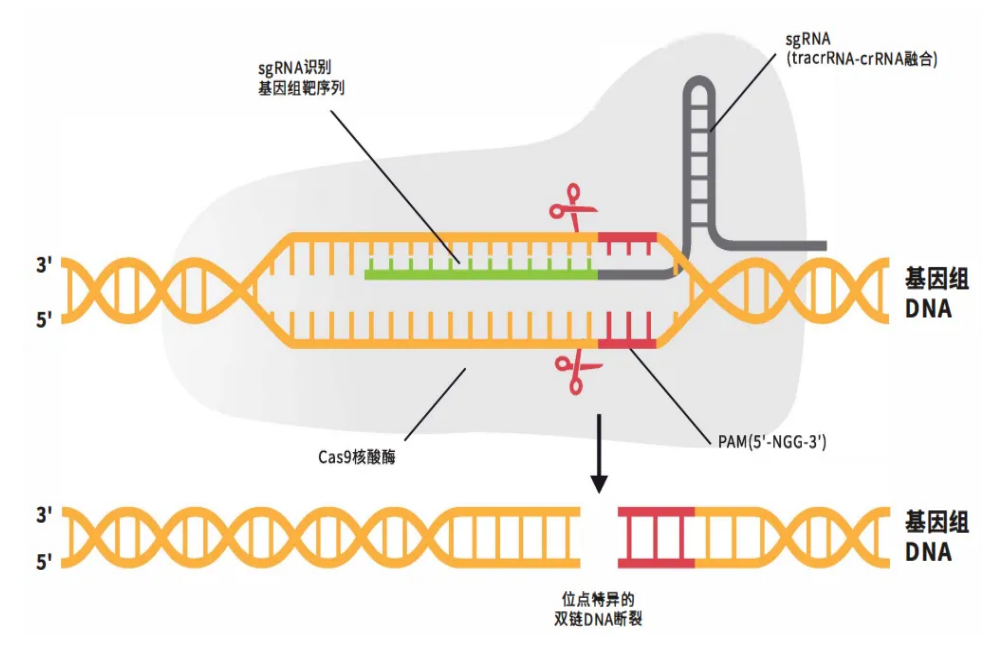
CRISPR/Cas9技術原理
切割后的DNA修復路徑如下:
路徑1:非同源末端連接(NHEJ),生物體內自發的SOS修復,修復期間會導致片段的插入或缺失。
路徑2:同源定向修復(HDR),引入高度同源的DNA修復模板時會啟動,實現片段的定向插入。
第三代技術相比于ZFNs和TALENs前兩代技術更有優勢,實驗設計難度降低、操作流程簡便、脫靶率低,細胞毒性低(如使用純蛋白RNP體系進行基因編輯)。但依舊有脫靶風險,當sgRNA與非目標序列部分匹配時,Cas9可能誤切關鍵基因(如癌癥相關基因)。不過基因編輯研發并未止步,2023年,CRISPR基因編輯藥物Casgevy上市,據2025年5月數據顯示90%以上的鐮狀細胞病患者實現“功能性治愈”;2024年,中國科學家更發現VII型CRISPR系統(Cas14),首次實現僅靶向RNA而不改變DNA序列的精準編輯,為基因治療開辟更安全的路徑。相信在未來CRISPR技術將更加精進,運用于個性化的精準醫療,造福人類。
隨著CRISPR技術從實驗室走向產業化,標準化工具鏈的完善成為推動臨床與農業應用的關鍵。針對基因編輯全流程的痛點——從Cas蛋白、sgRNA設計、Cas蛋白遞送到脫靶驗證,近岸蛋白已開發出覆蓋全鏈條的解決方案:
01 純蛋白編輯工具:Cas9(Cat.No.:E365)蛋白及其突變體,同時包含GMP級別的Cas9蛋白(Cat.No.:GMP-1701)、0脫靶的AaCas12bMax、enCas12Ultra等蛋白;
02 通用型一步法sgRNA制備:sgRNA體外轉錄試劑盒(Cat.No.:E399)在20μl體系下sgRNA產量可達10-30μg;
03 高效RNP遞送系統:首先在體外將sgRNA和Cas9蛋白混合成為sgRNA和Cas9的復合效應物,然后通過瞬時轉染技術(脂質體或電轉等)將復合效應物遞送到細胞內,進而實現對靶基因的編輯(相關實驗操作可后臺咨詢近岸蛋白公眾號);
04 脫靶驗證:T7核酸內切酶I有效檢測基因編輯之后形成的突變體,識別切割效率高。
三代基因編輯技術性能比對

以上便是基因編輯技術從ZFNs、TALENs到CRISPR/Cas9三代技術的進化史詩。而CRISPR革命的核心引擎——Cas蛋白家族,實則暗藏更精妙的分子江湖:下期我們將帶您深入CRISPR武器庫,解析不同Cas蛋白的PAM偏好、切割特性與應用場景,助您精準匹配研究需求。
作為深耕基因編輯領域的專業解決方案提供者,近岸蛋白將持續推出技術解析、實操指南與定制化工具,陪伴每一位科研工作者探索生命密碼的無限可能。敬請期待,下期再會!
相關產品推薦
CRISPR/Cas9相關產品
| 產品類型 | 優勢 | 目錄號 | 產品名稱 |
| RNA體外轉錄 | 通用型體外轉錄sgRNA | E399 | Universal sgRNA In Vitro one-step Transcription Kit |
| 一步法體外轉錄sgRNA | E369 | sgRNA In Vitro one-step Transcription Kit | |
| T7 RNA聚合酶轉錄 | E131 | T7 High Yield RNA Transcription kit | |
| SpCas9 | 通用型及突變體型 | GMP-1701 | Recombinant Cas9 Nuclease, GMP Grade |
| E365 | NLS-Cas9 Nuclease | ||
| E376 | NLS-Cas9 (D10A) Nickase | ||
| E377 | NLS-Cas9 (H840A) Nickase | ||
| E378 | NLS-Cas9 (D10A, H840A) Nuclease | ||
| E379 | NLS-Cas9-EGFP Nuclease | ||
| MR019 | Cas9 mRNA (N1-Me-Pseudo UTP) | ||
| MR107 | Cas9 mRNA | ||
|
AaCas12bMax |
零脫靶 | GMP-E375 | AaCas12bMax Enhanced, GMP Grade |
| E375 | AaCas12bMax Enhanced | ||
| MR205 | AaCas12bMax Enhanced mRNA (N1-Me-Pseudo UTP) | ||
| GR002 | AaCas12bMax Human TRAC sgRNA | ||
| GR003 | AaCas12bMax Human CIITA sgRNA | ||
| enCas12Ultra | 高編輯效率 | GMP-E393 | enCas12Ultra, GMP Grade |
| E393 | enCas12Ultra | ||
| MR208 | enCas12Ultra mRNA (N1-Me-Pseudo UTP) | ||
| piggyBac轉座子mRNA | 適用于動物和人細胞的轉座 | MR005 | piggyBac mRNA |
| 基因編輯效率檢測 | 80倍高保真性 | E035 | 2×Fast Pfu Master Mix |
| T7核酸內切酶I | M017 | T7 Endonuclease I | |
| DNA/RNA純化 | DNA純化磁珠 | N240 | NovoNGS® DNA Clean Beads |
| RNA純化磁珠 | N243 | RNA Clean Beads | |
| 病毒純化 | GMP級 | GMP-1707 | BenzoNuclease®, GMP Grade |
| GMP-1709 | BenzoNuclease® (Tag-free), GMP Grade | ||
| GMP-1711 | High Salt Active BenzoNuclease® (HSAB), GMP Grade |