iPSC神經再生療法:開啟神經修復的新紀元
隨著全球老齡化加劇,神經系統疾病的治療需求日益緊迫。由于人類中樞神經系統的再生能力極為有限,帕金森病、脊髓損傷等疾病的傳統治療手段往往難以實現根本性的修復。
誘導多能干細胞(iPSC)技術的成熟,為這一困境帶來了革命性解決方案。該技術通過導入特定轉錄因子,將體細胞重編程為具有多向分化潛能的干細胞,兼具無倫理爭議與低免疫排斥的獨特優勢。
其核心應用價值凸顯于三大方面:
•定向分化:可精準誘導為特定功能神經細胞(如多巴胺能神經元),用于替代治療。
•疾病建模:構建患者特異性疾病模型,成為高效藥物篩選的“先進試驗場”。
•臨床轉化:全球范圍內,從視網膜上皮移植到多巴胺祖細胞治療,多項臨床試驗已證實其巨大潛力。
iPSC神經再生療法,正將無數患者“無法治愈”的嘆息,轉化為生命重建的希望。
iPSC神經分化的精密調控網絡
iPSC向神經細胞的分化是一個高度精細和復雜的過程,需要通過多種細胞因子的協同作用,精確模擬體內神經發育的微環境。這一過程通常分為多個關鍵階段,每個階段都有特定的細胞因子發揮著不可或缺的調控作用。
1 神經誘導與模式化:奠定神經分化的基礎
iPSC神經分化的第一步是將其誘導為神經上皮細胞,這一過程需要抑制BMP/TGF-β信號通路。Activin A、Noggin(BMP拮抗劑)等細胞因子在此階段起著關鍵作用,它們能夠打破iPSC自我更新的狀態,驅動其向神經命運轉變。
接下來,新生成的神經上皮細胞需要進行特異性分化,以獲得目標細胞。以中腦多巴胺能神經元的分化為例,這一過程需要SHH、Wnt、FGF8等模式化因子的精確調控,模擬胚胎中腦發育的微環境。這些因子的濃度、活性穩定性及使用時序稍有偏差,就可能導致神經元身份的不準確,影響治療效果。
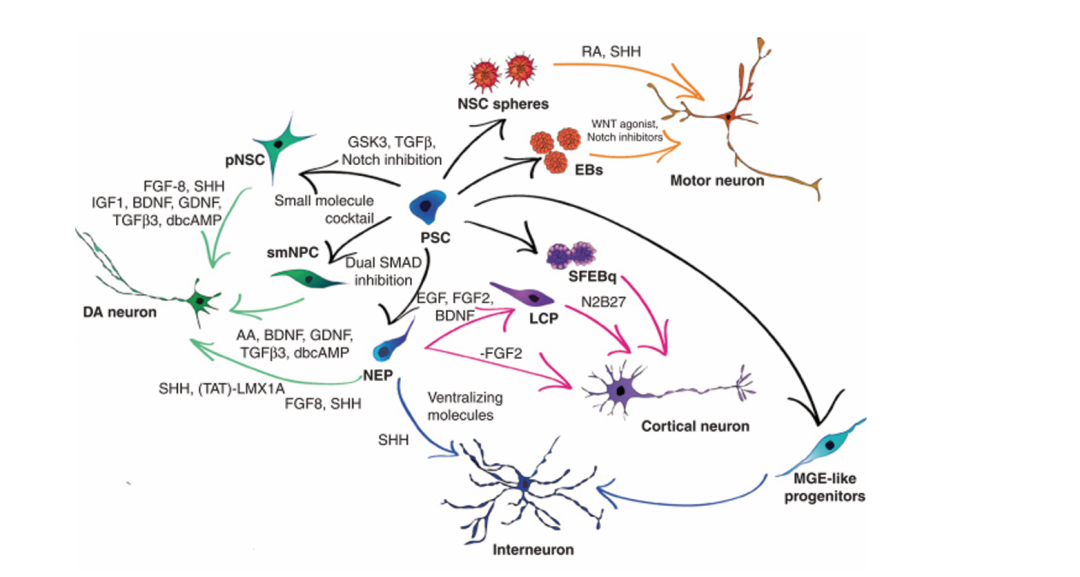
圖1.PSCs通過抑制策略先誘導為神經干細胞(NSC)球或中間前體細胞(如smNPC、NEP);后續經SHH、RA、FGF家族因子、小分子化合物等特異性因子誘導,可定向分化為多巴胺能(DA)神經元、運動神經元、皮質神經元、中間神經元等亞型,圖中同時標注了各階段對應的關鍵培養體系及信號調控分子[1]。
2 神經前體細胞的增殖與特化:確保細胞純度與方向
一旦獲得區域特異性的神經前體細胞,需要進一步促進其增殖和特化。研究表明,在脊髓神經前體細胞的分化過程中,通過精細的定向誘導,可獲得高純度(可達98%以上)的特定神經前體細胞,這些細胞表達神經祖細胞標志物(SOX2、PAX6)及區域特異性標志物(HOXB4)[2]。
此階段,FGF和EGF等因子被廣泛應用于加速神經前體細胞的增殖。通過優化因子的組合和濃度,研究人員能夠在體外獲得大量治療所需的目標細胞,為臨床應用提供充足的細胞來源。

圖2.人誘導多能干細胞(iPSC)向成熟運動神經元分化的流程示意圖。分化過程依次經歷Neural Rosettes形成、神經前體細胞(NPC)擴增、未成熟運動神經元階段,最終獲得成熟運動神經元[3]。
3 神經元成熟與功能整合:實現神經回路重建
獲得成熟的、有功能的神經元是iPSC神經再生治療的最終目標。在此階段,BDNF、GDNF等神經營養因子發揮著至關重要的作用。它們不僅支持神經元的存活和成熟,還促進突觸形成和神經遞質分泌,幫助新生的神經元整合到宿主神經網絡中。研究顯示,iPSC分化的運動神經元在BDNF、GDNF等因子作用下,能夠產生動作電位,這是神經細胞傳遞信號的關鍵能力[4]。
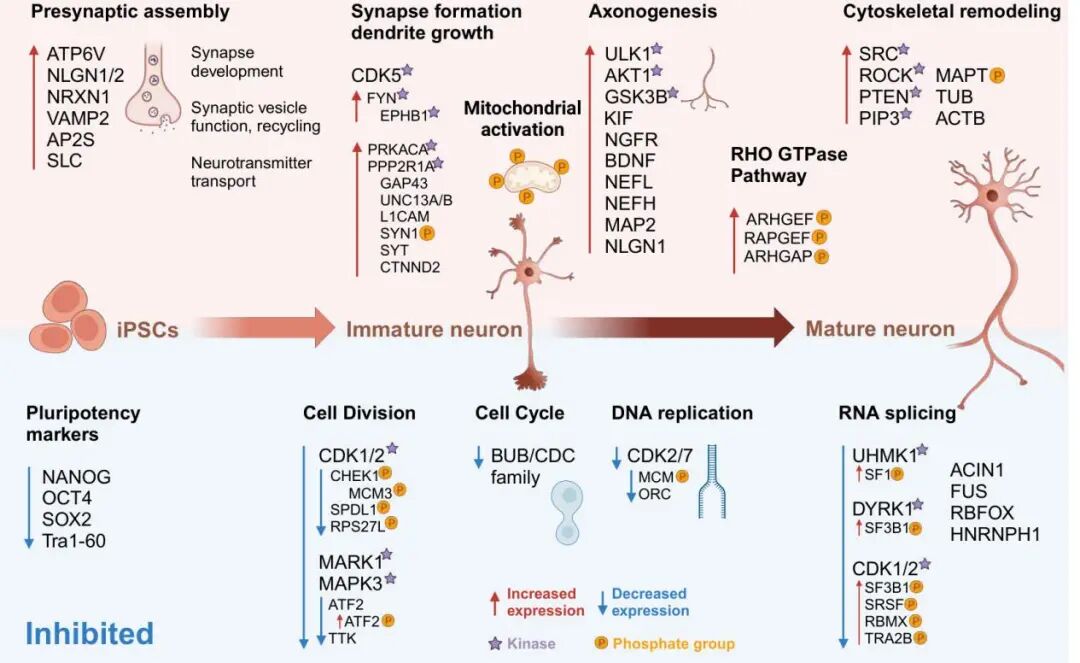
圖3.分化的三個階段(iPSCs→未成熟神經元→成熟神經元)對應的核心生物學過程:iPSCs階段多能性標志物(如NANOG、OCT4)表達受抑制;未成熟神經元階段涉及突觸組裝、線粒體激活等過程;成熟神經元階段則發生軸突發生、細胞骨架重塑等事件,同時標注了各過程對應的關鍵蛋白、激酶(*標注)及磷酸基團[5]。
4 細胞因子的質量控制:治療成功的關鍵保障
在iPSC神經分化過程中,細胞因子的質量直接影響分化結果的穩定性。
符合需求的GMP細胞因子需要具備以下特性:
- 高活性和純度:
確保分化路徑精準,減少非特異性效應; - 低宿主蛋白殘留:
降低免疫原性和非目標分化風險; - 卓越的批間一致性:
保證分化工藝的可重復性和穩定性。
近岸蛋白專注于提供符合客戶需求的GMP級別的細胞因子產品,包括Activin A、Noggin(BMP拮抗劑)、SHH、FGF8、BDNF、GDNF等,已助力客戶成功進行IND申報,為iPSC神經再生研究的標準化和產業化提供堅實支撐。
臨床進展與企業布局:iPSC神經再生治療的全球化發展
隨著iPSC神經分化技術的日益成熟,國內外多家研究機構和企業已在這一領域取得顯著進展,推動了iPSC神經再生療法從實驗室向臨床的轉化。
1 國內領軍企業的突破性進展
中國在iPSC神經再生領域的發展尤為迅速,多家企業已在特定細分方向取得全球首創的突破。
如士澤生物已在iPSC治療帕金森病和脊髓損傷方面取得多項里程碑進展。該公司開發的全球第一款異體通用"現貨型"iPSC衍生脊髓神經細胞新藥(XS228細胞注射液),已于2025年啟動治療脊髓損傷的注冊臨床I期研究,并成功完成全球首例受試者給藥。睿健醫藥、中盛溯源和躍賽生物等公司均在iPSC治療神經系統疾病領域有所布局,并正在加速推進臨床進展。呈諾醫學的ALF201是全球第一個獲批臨床的治療急性缺血性腦卒中的iPSC衍生細胞候選產品,已經進入臨床試驗階段。
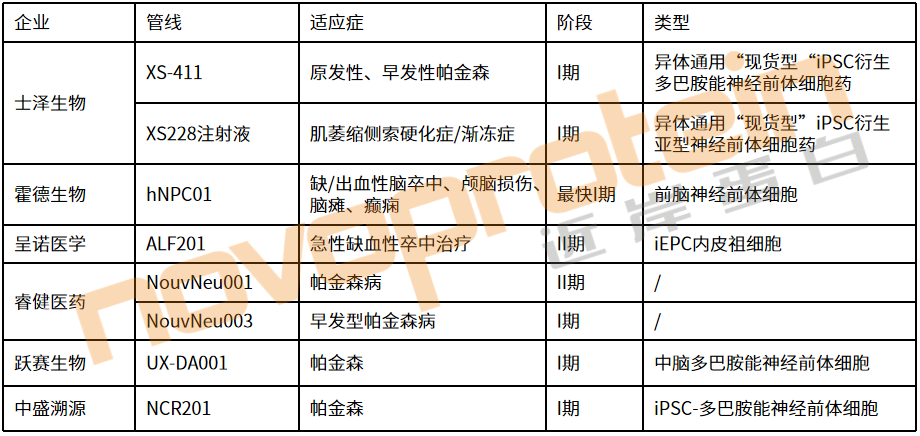
注:管線信息來源于官網,由近岸蛋白整理
如有紕漏,歡迎指正
2 國際研究進展與技術趨勢
在國際舞臺上,iPSC神經再生研究同樣如火如荼。拜耳公司治療帕金森病的在研產品BRT-DA01已進入臨床Ⅰ期,受試者均表現出良好的安全性和耐受性。近年來,新型治療策略不斷涌現。2025年,韓國建國大學與首爾大學研究團隊聯合開發了一種基于iPSC的新型神經再生治療技術,該技術以患者尿液細胞來源的iPSC分化出的神經嵴細胞為基礎,制備出攜帶神經生長因子(NGF)基因的細胞外囊泡(EV)。這種囊泡注射至周圍神經損傷小鼠模型后,顯著提高了受損區域的神經細胞存活率,抑制了炎癥反應,促進了軸突生長和再髓鞘化。值得注意的是,這種無需進行干細胞移植的治療方法規避了傳統干細胞治療可能帶來的腫瘤形成風險和免疫排斥問題,為神經再生治療提供了新思路[6]。
在疾病模型研究方面,科學家們將人類iPSC衍生的神經progenitors移植到小鼠大腦中,成功建立了人類神經網絡的體內模型。這一模型不僅能夠模擬人類神經網絡的發育和功能,還可用于研究Down syndrome等神經系統疾病的病理機制[7]。
iPSC神經再生的前景與展望
iPSC技術在神經再生領域的應用,代表了再生醫學的最前沿突破。隨著我們對神經分化機制的深入理解和細胞因子調控網絡的精確掌握,iPSC神經再生治療正逐步走向臨床現實。
從技術發展來看,iPSC神經再生領域正朝著標準化、精準化和產業化方向邁進。標準化制備的"現貨型"細胞制劑、精準靶向特定神經亞型的分化技術,以及符合國際標準的質量控制體系,共同構成了iPSC神經再生治療的堅實基礎。
盡管iPSC神經再生治療取得了顯著進展,但仍面臨一些技術挑戰,致瘤性風險是iPSC臨床應用中最為關注的安全問題之一。未完全分化的iPSC可能不正常增生形成腫瘤,而使用病毒載體進行重編程也可能因載體插入細胞基因組而誘發癌癥。
未來,隨著越來越多臨床試驗的開展和治療機制的闡明,iPSC神經再生療法有望在未來幾年內取得更大突破,為帕金森病、脊髓損傷、腦卒中等難治性神經系統疾病提供全新的治療選擇,最終實現神經功能的重建和患者生活質量的提升。
在iPSC神經再生治療的發展歷程中,高質量的細胞因子作為精準調控分化過程的核心要素,將繼續發揮著不可或缺的作用。
在iPSC領域,近岸蛋白提供RUO級和GMP級基質蛋白(Vitronectin/Laminins)、細胞因子、Cas9/Cas12b蛋白、全能核酸酶以及mRNA原料酶等CGT關鍵生產原料,加速iPSC客戶藥物研發進程。
產品數據
高活性基質蛋白支持iPSC穩定傳代和干性維持

iPSC分別在Vitronectin、Laminin511和Laminin521的支持培養下,進行穩定連續傳代培養P1至P5,均可維持良好的生長,細胞形態均一正常。
有效維持細胞干性
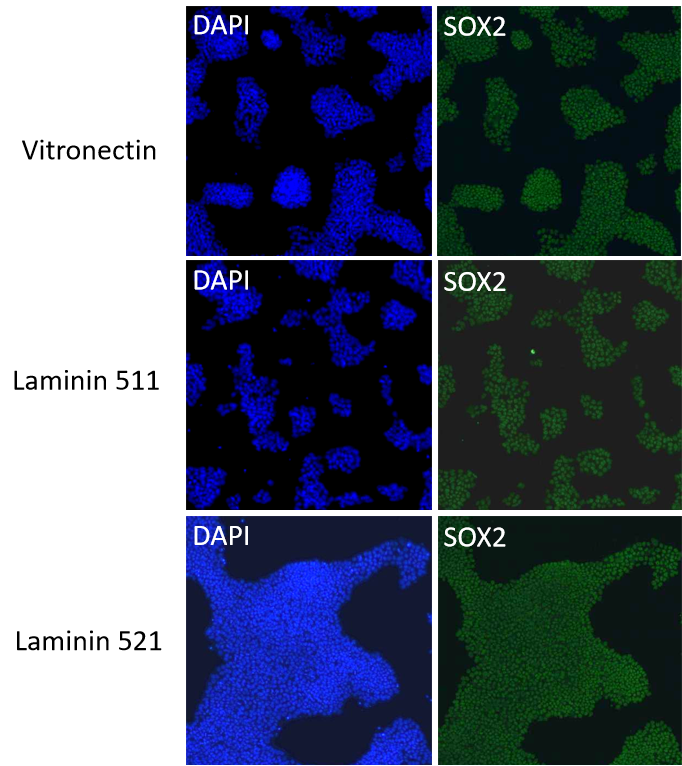
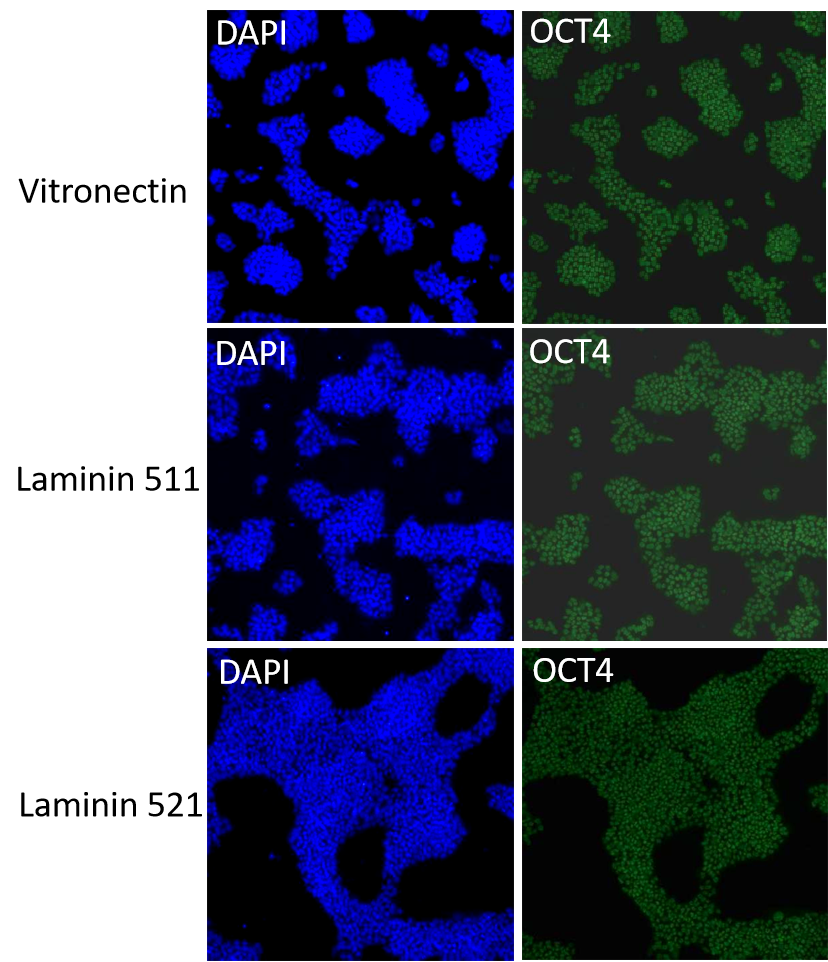
Vitronectin、Laminin511和Laminin521培養的iPSC可以有效維持干性相關轉錄因子的表達。
高活性iPSC誘導分化細胞因子
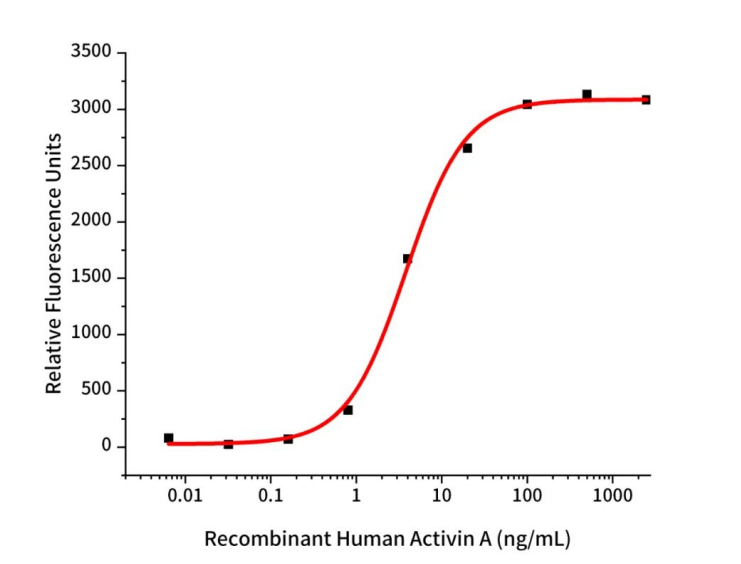
Recombinant Human Activin A (Cat.No.:GMP-C687)
Measured by its ability to induce SMAD signaling in 293-Activin A Res cells. The specific activity of recombinant human Activin A is ≥1.0 x 103 IU/mg, which is calibrated against the human Activin A Standard (NIBSC code: 91/626).
相關產品
| 應用場景 | 主要產品 |
| iPSC衍生的T細胞 | bFGF、BMP-4、FLT3L、IL-3、IL-7、SCF、VEGF165 |
| iPSC衍生的NK細胞 | BMP-4、FLT3L、IL-2、1L-3、IL-7、IL-15、SCF、VEGF165 |
| iPSC衍生的巨噬細胞 | bFGF、BMP-4、FLT3L、GM-CSF、IL-3、IL-7、 LR3 IGF-1、M-CSF、SCF、VEGF165 |
| iPSC衍生的多巴胺能神經元 | bFGF、BDNF、FGF8b、GDNF、Noggin、SHH、TGF-beta3 |
| iPSC衍生的神經細胞 | bFGF、BDNF、CNTF、GDNF、LR3 IGF-1、SHH、PDGF-BB、FGF-10 |
| iPSC衍生的星形膠質細胞 | ActivinA、BMP-4、bFGF、CNTF、EGF、LR3 IGF-1、NRGl-Beta |
| iPSC衍生的少突膠質細胞 | HGF、LR3 IGF-1、NT-3、PDGF-AA |
| iPSC衍生的小膠質細胞 | CD200、CX3CL1、IL-34、M-CSF、TGF-beta 1 |
| iPSC衍生的神經祖細胞 | bFGF、NT-3 |
| iPSC衍生的心肌細胞 | Activin A、BMP-4、bFGF、DKKl、SCF、TGF-beta l、Transferrin、VEGF165、Wnt 3a |
| iPSC衍生的胰島細胞 | ActivinA、BTC、bFGF、EGF、KGF |
| iPSC衍生的角膜細胞 | IGF-1、LIF、IL-6、IL-11、TNF-α |
| iPSC基質蛋白 | Vitronectin、Laminin521、Laminin511 |
| 基因編輯 | Cas9、AsCas12bMAX、enCas12hf、enCas12Ultra |
| 核酸消除 | 全能核酸酶及殘留檢測試劑盒 |
參考文獻
[1]De Corti, M., (2015). Schéma des différents protocoles existants pour différentier des cellules souches embryonnaires ou induites en différents sous-types de neurones. In Stem Cell Differentiation Protocols: Neuronal Lineages (pp. [69]). Springer.
[2]Shi, Z., De Corti, M., (2025). Human iPSC-derived spinal neural progenitors enhance sensorimotor recovery in spinal cord-injured NOD-SCID mice via differentiation and microenvironment regulation. Stem Cell Reports.
[3]High Efficiency iPSC Reprogramming for Regenerative Medicine: RNA-LNP Complete Kits. 技術資料,2023.
[4]Lara, E., Hao, Y., Li, Z., et al. (2024). Functional maturation of human iPSC-derived motor neurons requires BDNF/GDNF-mediated synaptic integration and action potential generation. Acta Neuropathologica Communications, 14(1), 178.
[5]Hao, Y., Li, Z., Lara, E., et al. (2025). Temporal dynamics of proteome and phosphorproteome during neuronal differentiation in the reference KOLF2.1J iPSC line [Preprint]. Sciety.
[6]Cho, S. G., Yeo, H. C., Seok, J. K., Kim, Y. J., Jang, S. B., Lee, N. H., ... & Lee, S. W. (2025). NGF-enriched extracellular vesicles derived from urine-derived iPSC neural crest cells promote peripheral nerve regeneration. Acta Neuropathologica Communications, 13, 125.
[7]Real, R., Peter, M., Trabalza, A., Khan, S., Smith, M. A., Dopp, J., ... & De Paola, V. (2018). In vivo modeling of human neuron dynamics and Down syndrome. Science, 362(6416), eaau1810.