IF=6.2!中科院茶葉研究所林智團隊突破性成果:非靶向特定修飾代謝組學首次系統解析植物糖基化代謝物

英文標題:Nontargeted Modification-Specific Metabolomics Investigation of Glycosylated Secondary Metabolites in Tea (Camellia sinensis L.) Based on Liquid Chromatography-High-Resolution Mass
中文標題:基于液相色譜-高分辨質譜的茶糖基化次級代謝產物非靶向特定修飾代謝組學研究
發表期刊:Journal of agricultural and food chemistry
影響因子:6.2
研究背景
內源性修飾(如糖基化和酰化)廣泛存在于生物體中,調控多種生物過程,并在植物小分子代謝物中尤為常見。這些修飾通過改變代謝物的極性、穩定性、生物活性等,幫助植物抵御生物和非生物脅迫,同時生成復雜的次級代謝物(如抗氧化劑、輔酶及輻射防護分子)。糖基化代謝物在植物中具有高度物種特異性,是藥物、營養和風味成分的重要來源,但其大規模研究受限于分析技術。
近年來,代謝組學與高分辨質譜技術的進步推動了修飾代謝物的系統解析。通過引入非靶向特定修飾代謝組學(結合源內碰撞誘導解離技術),研究者已實現對尿液中900余個修飾代謝物(如乙酰化、糖基化等)的高通量檢測,并發現大量未收錄于數據庫的新化合物。
茶 (Camellia sinensis)作為全球第二大飲品,其健康效應與風味特征主要源于豐富的次級代謝物(如黃酮類、類黃酮糖苷等)。然而,茶中仍有大量糖基化代謝物未被發現。本研究首次應用上述方法,系統解析茶中葡萄糖基化、半乳糖基化、鼠李糖基化、蕓香糖基化及藤黃糖基化等修飾的次級代謝物,為茶的代謝組復雜性及功能成分挖掘提供新視角。
研究結果
代謝物糖基化鑒定策略
如圖1所示,內源性代謝物糖基化會為代謝物底物增加特定分子量。在源內碰撞誘導解離(ISCID)過程中,通過中性丟失路徑可去除所添加的配體,從而產生特定的中性丟失模式。在茶中糖基化代謝物的初步分析中,以槲皮素3-葡萄糖苷、山柰酚3-半乳糖苷、楊梅素3-鼠李糖苷、槲皮素3-蕓香糖苷和水楊酸甲酯藤黃糖苷為標準品,分別研究了葡萄糖基化、半乳糖基化、鼠李糖基化、蕓香糖基化和藤黃糖基化代謝物的特異性中性丟失(糖基化代謝物結構見圖2)。
如圖3所示,葡萄糖基化/半乳糖基化、鼠李糖基化、蕓香糖基化和藤黃糖基化修飾的代謝物分別表現出理論特征中性丟失(NL)值為162.0528(C?H??O?)、146.0579(C?H??O?)、308.1107(C??H??O?)和294.0951(C??H??O?),這些特征可作為識別特定糖基化的指標(表1)。值得注意的是,葡萄糖基化和半乳糖基化的NL值均為162.0528,僅通過NL值無法區分兩者。因此,需引入色譜保留行為:在C??色譜柱上,半乳糖基化結合物的洗脫時間早于葡萄糖基化結合物。借助高分辨Q-TOF/MS儀器的優勢,可輕松區分葡萄糖基化/半乳糖基化(NL=162.0528)與咖啡酰化(NL=162.0317),以及鼠李糖基化(NL=146.0579)與香豆酰化(NL=146.0368),而傳統三重四極桿或Q-trap儀器的中性丟失掃描無法實現此區分。需注意的是,若化合物的糖基部分進一步發生酰化(如沒食子酰化、香豆酰化或咖啡酰化),則通過ISCID無法產生對應的糖基化特征NL,因此無法通過非靶向特定修飾代謝組學方法檢測到此類化合物。
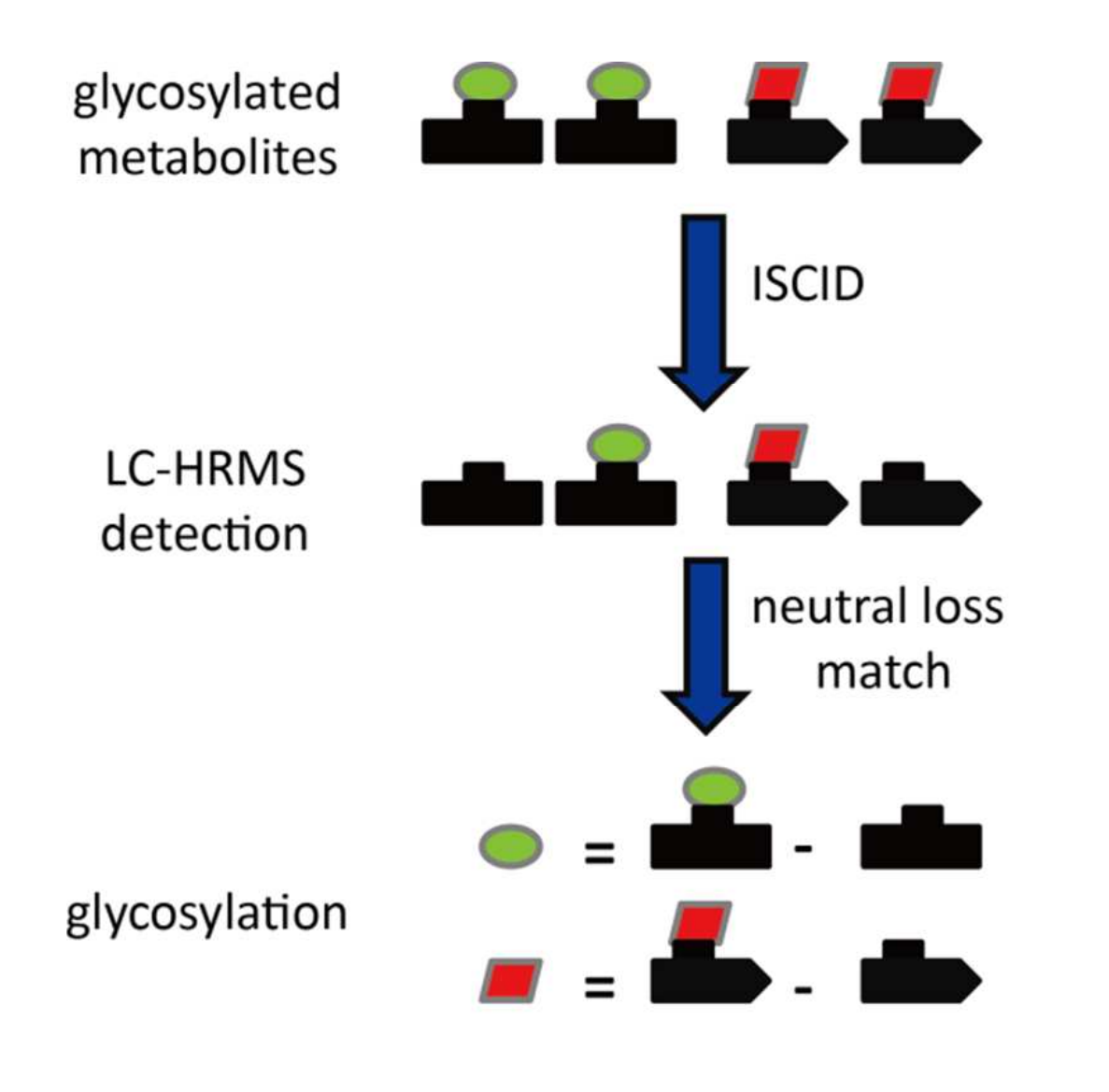
圖1 茶中代謝物糖基化映射策略
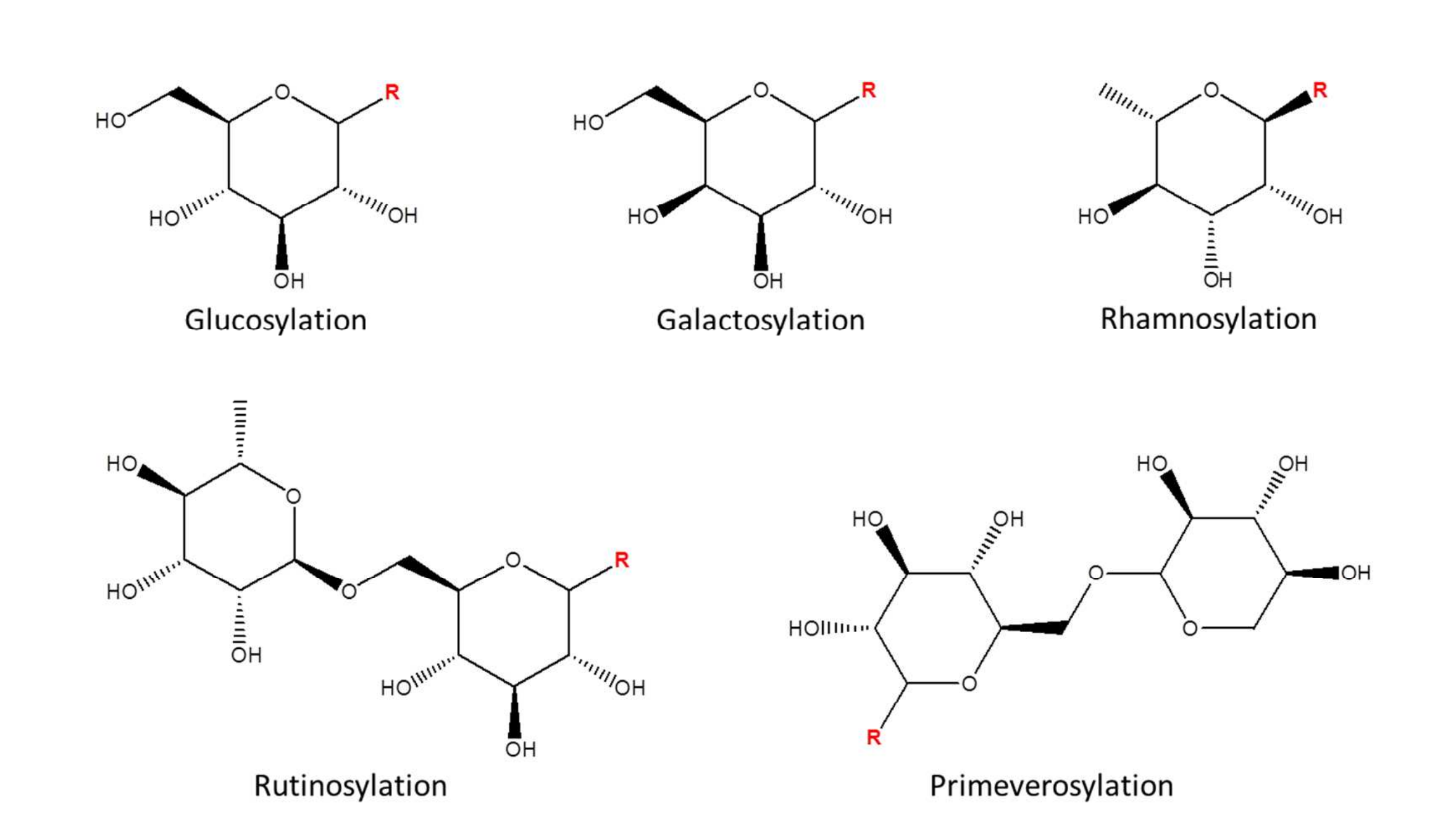
圖2 本研究涵蓋的代謝物糖基化結構
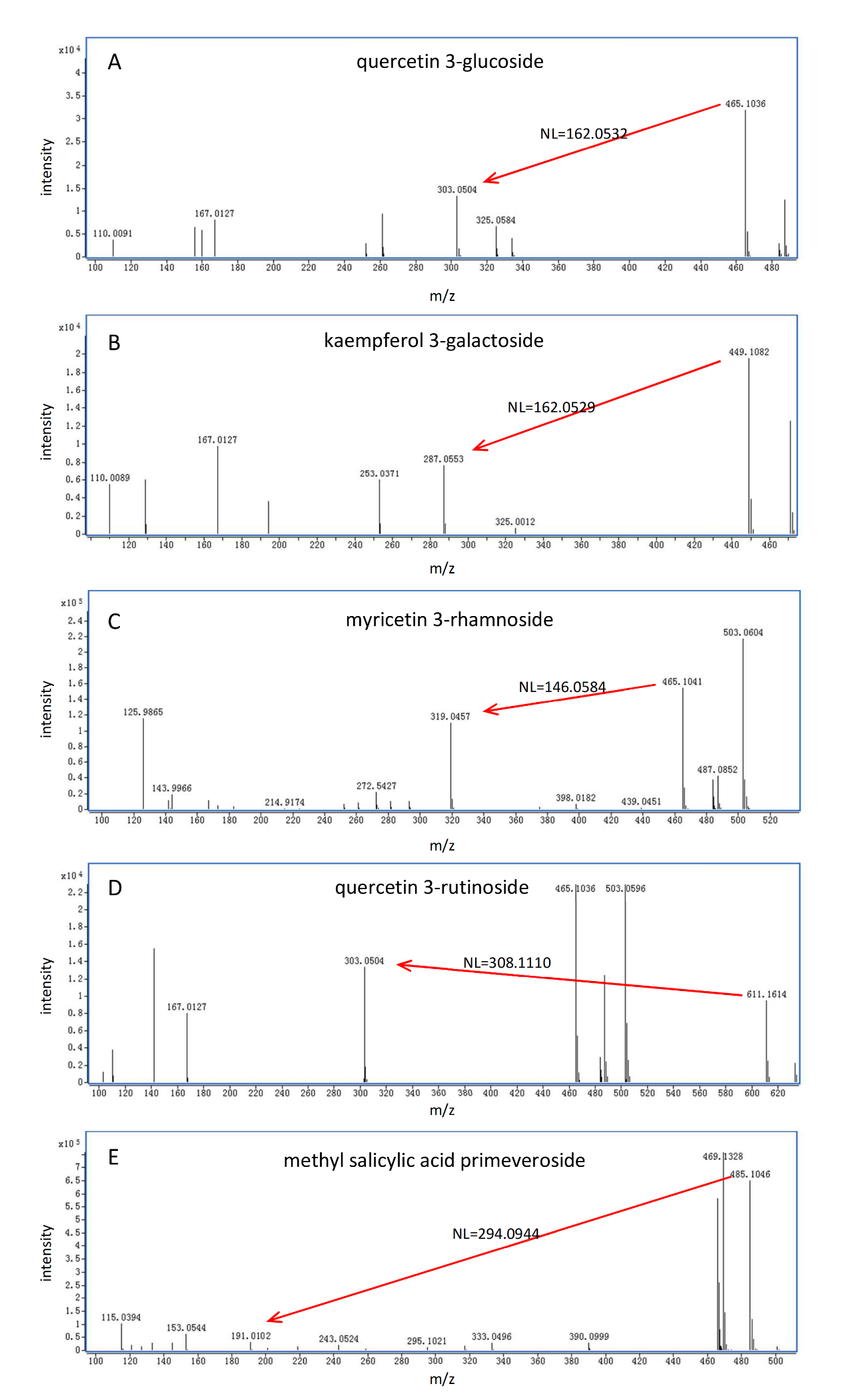
圖3 糖基化代謝物的典型ISCID碎片模式
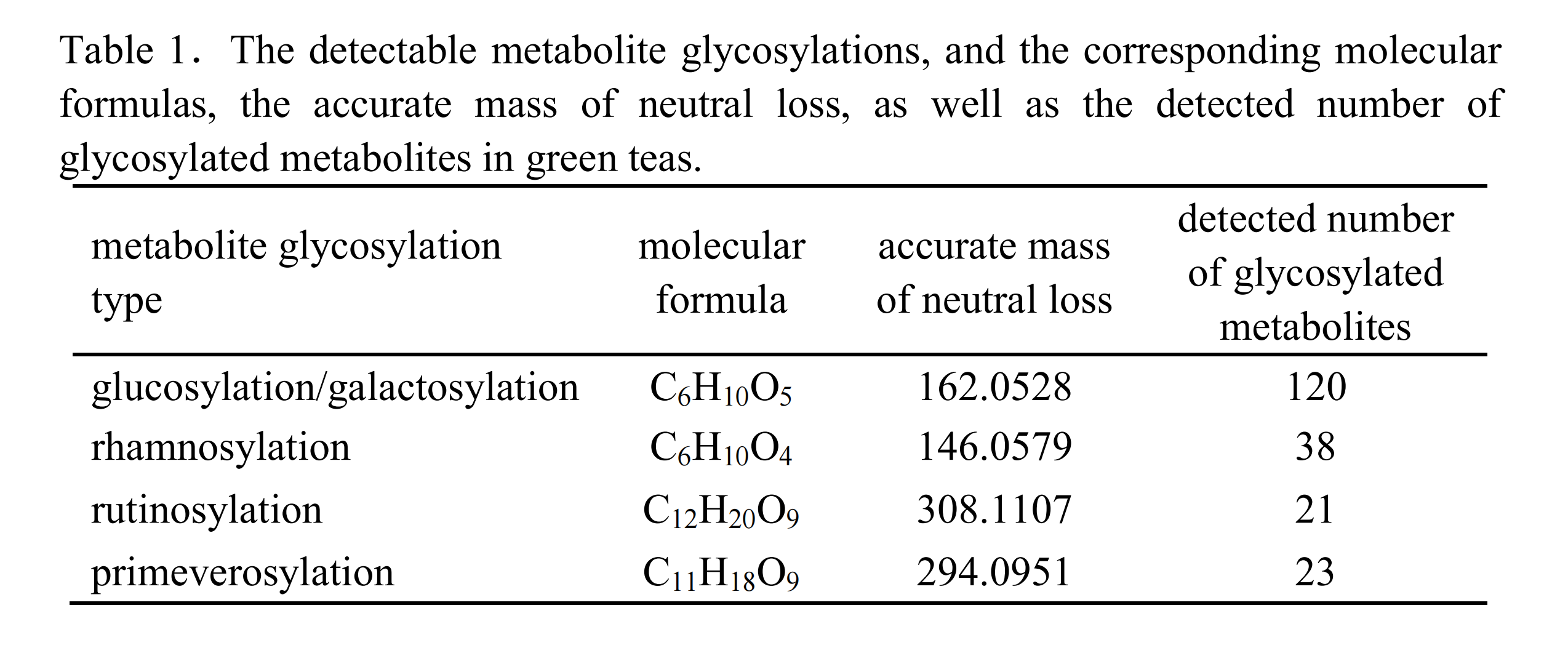
表1 可檢測的代謝物糖基化和相應的分子式,中性損失的準確質量,以及檢測到的綠茶中糖基化代謝物的數量
綠茶糖基化代謝物的分析
14種茶樹品種混合的綠茶樣本中,共鑒定出202種化合物,包括120種葡萄糖基化/半乳糖基化代謝物、38種鼠李糖基化代謝物、21種蕓香糖基化代謝物和23種藤黃糖基化代謝物(表1)。隨后,通過碰撞誘導解離(CID)模式下的串聯質譜進一步驗證代謝物的糖基化類型。其中144種化合物表現出源自特定糖基基團的中性丟失(表S1)。剩余的58種化合物因豐度較低,可能在MS²譜中未能檢測到目標特征,因此未被進一步分析。上述144種化合物被選為后續結構鑒定的對象。
輔助糖基化代謝物的鑒定
得益于液相色譜-質譜聯用技術(LC-MS)靈敏度和分辨率的快速提升,單個植物樣本(如綠茶浸提物)中可同時獲取數百甚至上千個離子特征信號。然而,僅極少數色譜和質譜信號能被結構解析,這一限制已成為植物代謝組學研究的瓶頸。
在本研究中,144種通過CID-MS²驗證的糖基化化合物中,有61種可通過TMDB和HMDB數據庫檢索實現結構解析(表S1)。然而,糖基化代謝物的結構異構體和立體異構體無法僅通過串聯質譜完全鑒定,且部分離子特征可能匹配多個糖基化代謝物候選物。因此,優先選擇茶中已報道的候選物,并通過標準品驗證(12種已通過標準品確認)進一步驗證。這表明,非靶向特定修飾代謝組學方法可有效解析茶中的糖基化代謝物。
通過提前解析糖基部分,非靶向特定修飾代謝組學策略能克服代謝物鑒定的局限性,即大量糖基化代謝物尚未被發現或收錄于數據庫。該方法的策略如下(圖4):
將推測糖基化代謝物列表中的離子特征導入代謝組學數據庫進行結構鑒定;
若未匹配到候選物(0 hit),則將對應底物([M-sugar]+)作為替代目標進行數據庫檢索;
通過結合底物部分和糖基部分解析離子特征的結構。(注:此處的“底物”定義為優先選擇茶中已報道的化合物作為參考)。
應用此策略,在綠茶浸提物中額外鑒定了40種新糖基化代謝物(表S1)。
以茶氨酸葡萄糖苷的鑒定為例:①一個離子特征(m/z=337.1599,保留時間2.26min)在非靶向特定修飾代謝組學分析中表現出NL=162.0524(對應葡萄糖基),因此被假設為糖基化代謝物;②該離子特征在TMDB和HMDB數據庫中未匹配到候選物;③通過提前解析葡萄糖基,將其底物(m/z=175.1071,丟失一個葡萄糖)檢索HMDB數據庫,匹配到茶氨酸(茶中特有游離氨基酸,占干茶重量的1-2%)。因此,該離子特征被鑒定為茶氨酸葡萄糖苷(1-茶酰葡萄糖),并通過合成標準品進一步確認(圖5)。由于該化合物此前未被報道,未來需開展大量工作以驗證其他新糖基化代謝物。
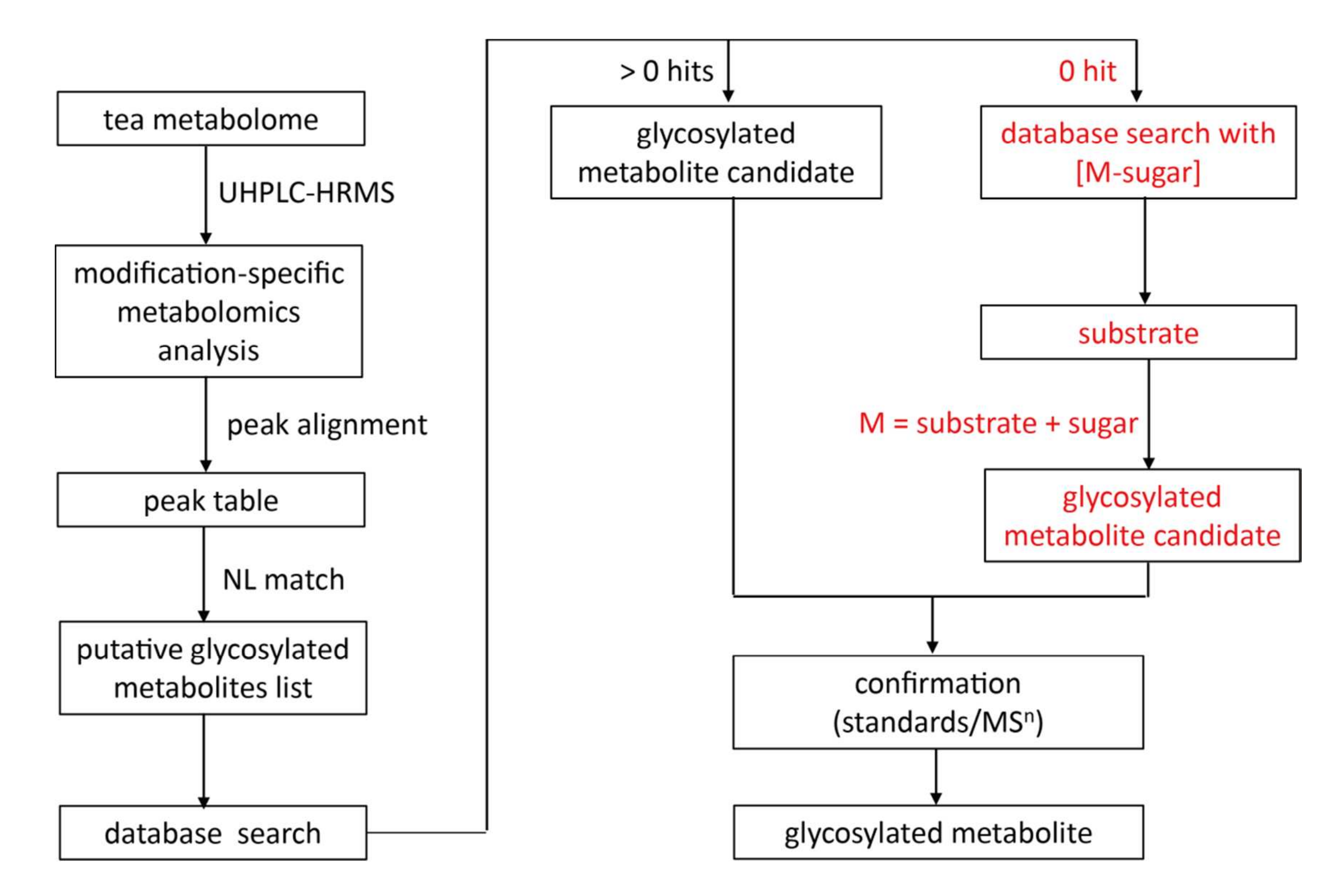
圖4 茶樹中糖基化代謝物發現與鑒定的特定修飾代謝組學流程
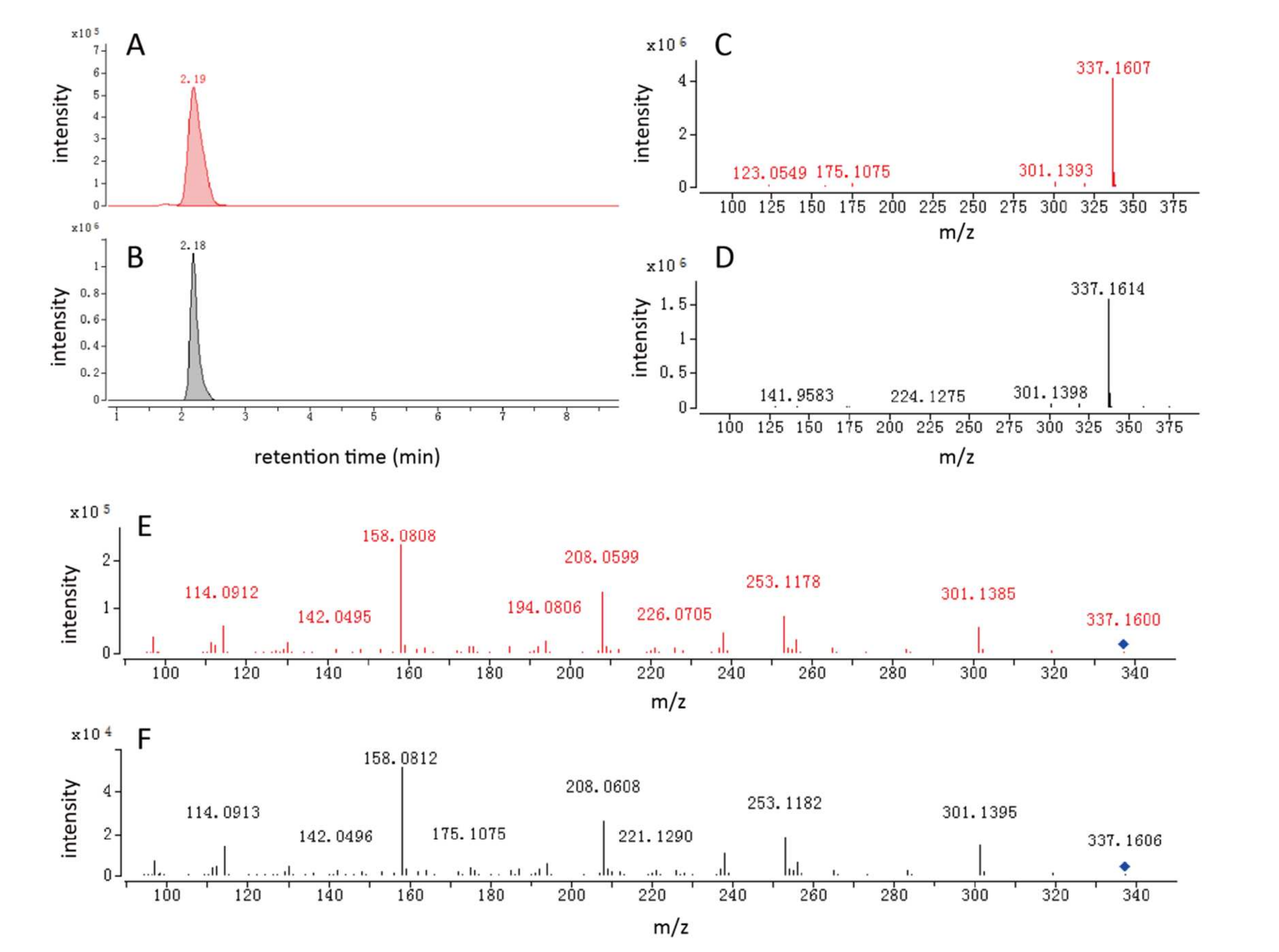
圖5 茶氨酸葡萄糖苷的提取離子色譜圖與質譜圖
不同品種綠茶中糖基化代謝物的差異
作為概念驗證,非靶向特定修飾代謝組學被應用于發現不同茶樹品種間主要差異的糖基化代謝物;14個品種中糖基化代謝物的相對定量結果見表S2。如圖6A所示,14種綠茶混合樣本(QC樣本)在主成分分析(PCA)得分圖中心密集聚集,表明代謝組學分析具有良好的重復性。Zhenghedabai、Fuzao 2、Maoxie、Longjing 43、Yuemingxiang、Xicha 5和Echa 1的糖基化代謝物譜相似,這些品種通常用于制作非發酵茶(Fuzao 2、Longjing 43、Xicha 5、Echa 1)、輕發酵茶(Zhenghedabai)和半發酵茶(Maoxie、Yuemingxiang)。另一方面,Jianbohuang、Ningzhou 2、Gaoyaqi、Zhuyeqi、Wannong 95、Huang Guanyin和Fuyun 6的譜相似,這些品種通常用于制作發酵茶(Jianbohuang、Ningzhou 2、Gaoyaqi、Zhuyeqi、Wannong 95、Fuyun 6)和半發酵茶(Huang Guanyin)。
通過PCA載荷圖(圖6B)篩選出對茶品種差異起主要作用的糖基化代謝物。如圖7所示,木犀草素-3-半乳糖苷、槲皮素-3-半乳糖苷、槲皮素-3-半乳糖基蕓香糖苷、山柰酚-3-半乳糖苷和山柰酚-3,3’-二半乳糖苷在PCA得分圖左側區域(Zhenghedabai等品種)顯著高于右側區域(Jianbohuang等品種),而山柰酚-3-葡萄糖苷、山柰酚-3-蕓香糖苷、山柰酚-3-葡萄糖基蕓香糖苷、N’-甲酰犬尿氨酸二葡萄糖苷和麥拉三糖在左側區域顯著低于右側區域。這表明,Zhenghedabai等品種的代謝物半乳糖基化活性更旺盛,而Jianbohuang等品種的葡萄糖基化活性較低。因此,推測糖基化/半乳糖基化比例可能與茶葉加工適配性相關:高半乳糖基化水平與非發酵茶加工適配性正相關,而高葡萄糖基化水平與發酵茶加工適配性正相關。
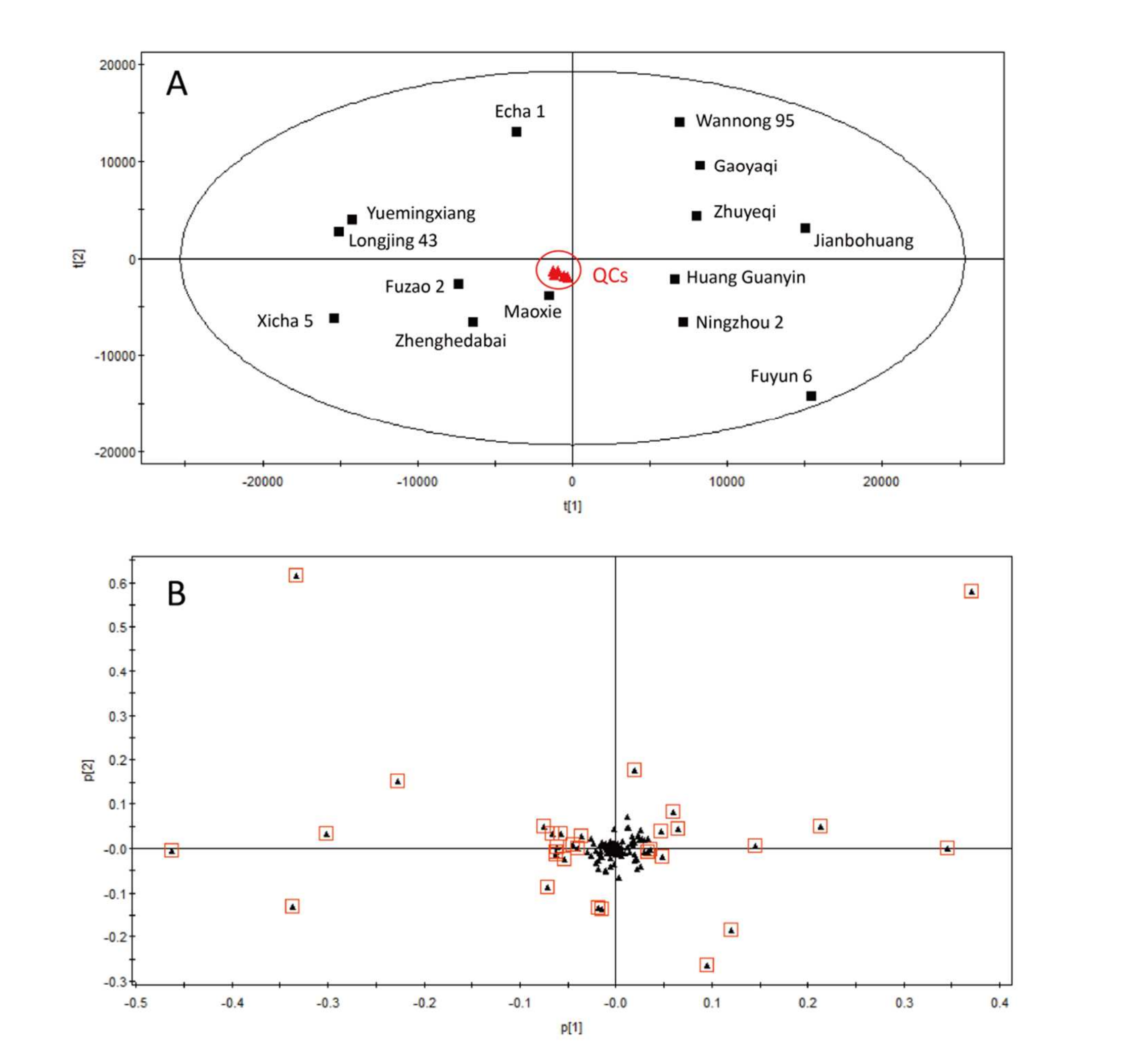
圖6 十四種綠茶品種樣本的主成分分析(PCA)
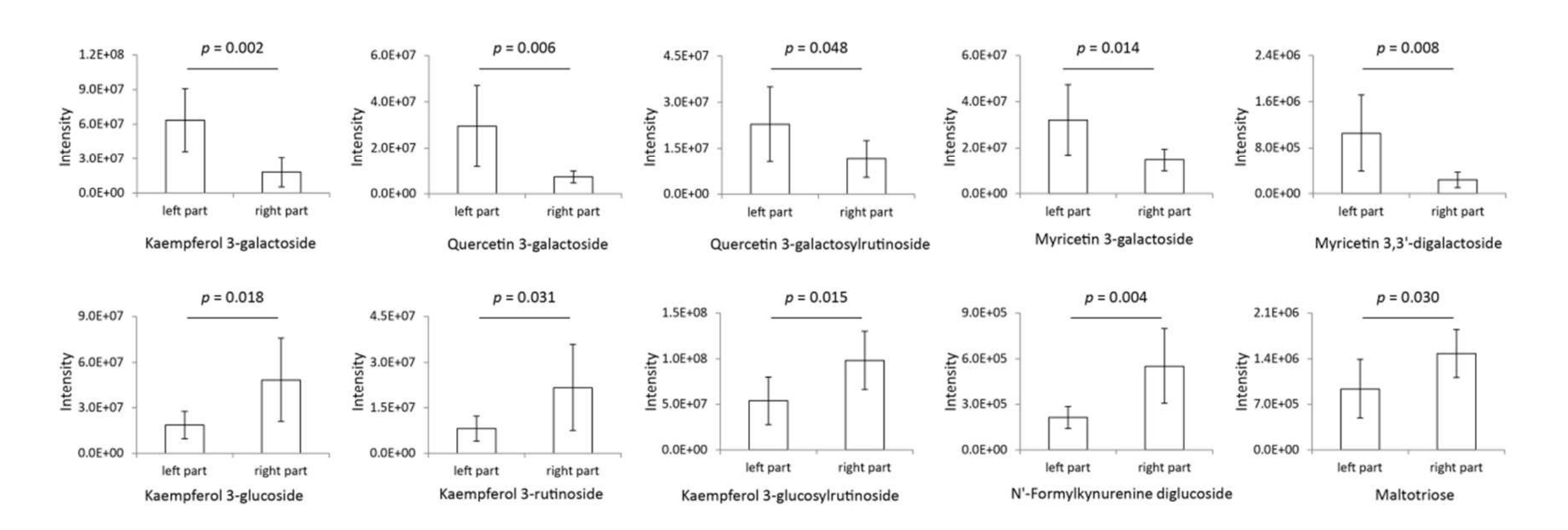
圖7 PCA得分圖左右區域品種間的主要顯著糖基化代謝物差異
研究結論
本研究成功應用非靶向特定修飾代謝組學方法,系統解析了綠茶中葡萄糖基化、半乳糖基化、鼠李糖基化、蕓香糖基化和素糖基化次級代謝物的特征。在綠茶中的應用中,總計檢測到120種葡萄糖基化/半乳糖基化代謝物、38種鼠李糖基化代謝物、21種蕓香糖基化代謝物和23種素糖基化代謝物。其中,61種糖基化代謝物可通過現有茶代謝物數據庫初步鑒定。此外,得益于糖基部分的預先注釋,該方法顯著輔助了代謝物鑒定,額外發現了40種新型糖基化代謝物。總的來說,該方法顯著擴大了糖基化代謝物的覆蓋范圍,并提升了未知代謝物的結構鑒定能力,為植物中新型糖基化代謝物的發現與鑒定提供了可行策略。未來可進一步擴展至其他重要修飾(如鞣酸化、肉桂酰化、香豆酰化和咖啡酰化)的植物代謝組研究。