
期刊:Nature communications
影響因子:14.7
主要技術(shù):單細(xì)胞RNA測序
導(dǎo)語
端腦的神經(jīng)活動主要由源自胚胎神經(jīng)節(jié)隆起(GE)的皮質(zhì)GABA能神經(jīng)元和皮質(zhì)下區(qū)GABA能神經(jīng)元塑造。然而,由于神經(jīng)元亞型高度復(fù)雜,人們對這些神經(jīng)元分化的機(jī)制仍知之甚少。本研究結(jié)合單細(xì)胞RNA測序和基因功能缺失實驗,發(fā)現(xiàn)小鼠中Foxg1基因的缺失會導(dǎo)致皮質(zhì)神經(jīng)元分化為皮質(zhì)下區(qū)神經(jīng)元。解析了神經(jīng)節(jié)隆起譜系的發(fā)育軌跡,揭示了FOXG1驅(qū)動的轉(zhuǎn)錄程序如何決定不同譜系中的神經(jīng)元亞型分化,并鑒定了調(diào)控譜系分化的關(guān)鍵轉(zhuǎn)錄因子。這項研究不僅闡明了皮質(zhì)和皮質(zhì)下區(qū)域神經(jīng)元群體生成的調(diào)控機(jī)制,還為GABA能神經(jīng)元相關(guān)疾病的發(fā)病機(jī)制提供了轉(zhuǎn)錄組學(xué)層面的新見解。
主要技術(shù)
單細(xì)胞RNA測序
(技術(shù)服務(wù)由伯豪生物提供)
研究結(jié)果
1. 神經(jīng)節(jié)隆起條件性敲除Foxg1驅(qū)動GABA能神經(jīng)元向皮質(zhì)遷移
為探究GE來源的GABA能神經(jīng)元的發(fā)育機(jī)制。作者通過條件性敲除Foxg1,發(fā)現(xiàn)Foxg1缺失導(dǎo)致GABA能神經(jīng)元在E16.5和E18.5階段異常聚集在皮質(zhì)下區(qū),而在大腦皮質(zhì)和嗅球(OB)中的分布顯著減少,表明Foxg1缺失影響了神經(jīng)元從GE向皮質(zhì)和嗅球的遷移。單細(xì)胞RNA測序分析E16.5階段的GE細(xì)胞群,發(fā)現(xiàn)Foxg1條件性敲除導(dǎo)致MGE來源的LHX6high皮質(zhì)中間神經(jīng)元(CINs)比例顯著降低,而紋狀體中間神經(jīng)元(SINs)和蒼白球典型神經(jīng)元比例增加。免疫染色證實了SST+中間神經(jīng)元在腹側(cè)前腦異常聚集。研究還意外發(fā)現(xiàn)一類表達(dá)NKX2-1的紋狀體投射神經(jīng)元(SPNs)新亞型,具有典型的SPN分子特征(Meis2、Foxp1/2高表達(dá)),并通過譜系追蹤實驗證實這些神經(jīng)元確實來源于MGE而非傳統(tǒng)認(rèn)為的腹側(cè)LGE。
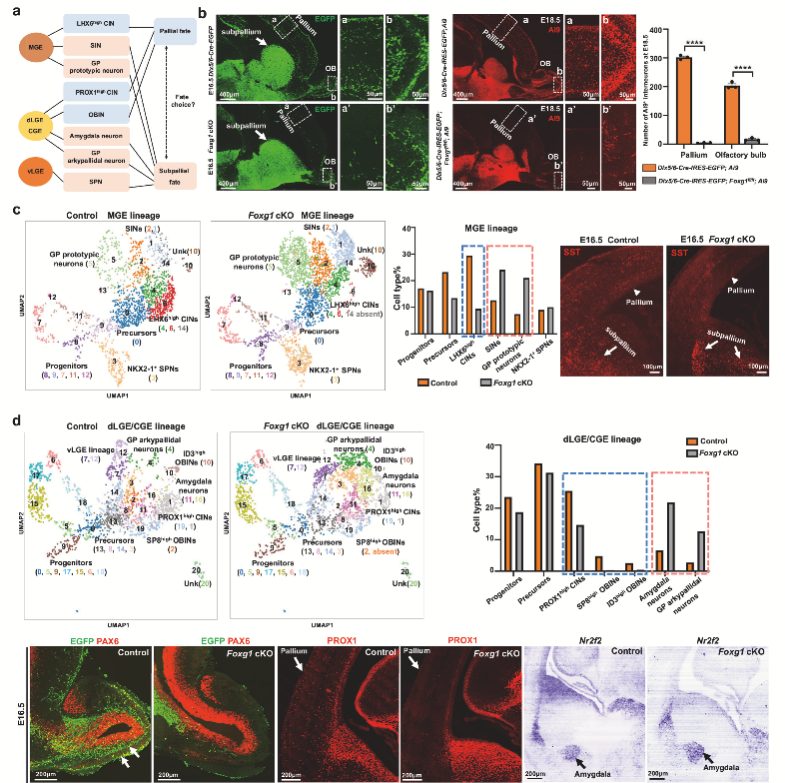
圖1 神經(jīng)節(jié)隆起中,F(xiàn)oxg1缺失驅(qū)動GABA能神經(jīng)元向皮質(zhì)遷移
2. LHX6high CIN 向蒼白球的轉(zhuǎn)化開始于 Foxg1cKO MGE 譜系的前體階段
為深入解析Foxg1敲除對MGE譜系神經(jīng)元的影響,作者對單細(xì)胞數(shù)據(jù)做了進(jìn)一步分析。RNA速率分析揭示了對照組MGE神經(jīng)元譜系兩條主要發(fā)育軌跡:一條從祖細(xì)胞經(jīng)共同前體狀態(tài)分化為LHX6high CINs、SINs和GP典型神經(jīng)元;另一條則較早地從祖細(xì)胞分化為NKX2-1+ SPNs。Foxg1敲除導(dǎo)致LHX6high CINs的發(fā)育軌跡明顯向SINs和GP神經(jīng)元方向偏移。共同前體細(xì)胞(C0)再聚類分析,發(fā)現(xiàn)Foxg1敲除導(dǎo)致前體細(xì)胞的轉(zhuǎn)錄因子表達(dá)譜發(fā)生顯著改變:促CIN分化的Zeb2和Cux2下調(diào),而促腹側(cè)神經(jīng)元分化的Nkx2-1和Lhx8上調(diào)。Nkx2-1下游靶標(biāo)Lhx6在前體細(xì)胞中表達(dá)升高,暗示腹側(cè)神經(jīng)元命運增強。定量分析顯示,F(xiàn)oxg1敲除小鼠中pre-CINs比例急劇下降,而pre-SINs/pre-GP神經(jīng)元比例顯著增加。以上結(jié)果表明,F(xiàn)oxg1缺失通過改變前體細(xì)胞的轉(zhuǎn)錄程序,促使MGE譜系神經(jīng)元從皮質(zhì)CINs向腹側(cè)SINs和GP神經(jīng)元轉(zhuǎn)變。
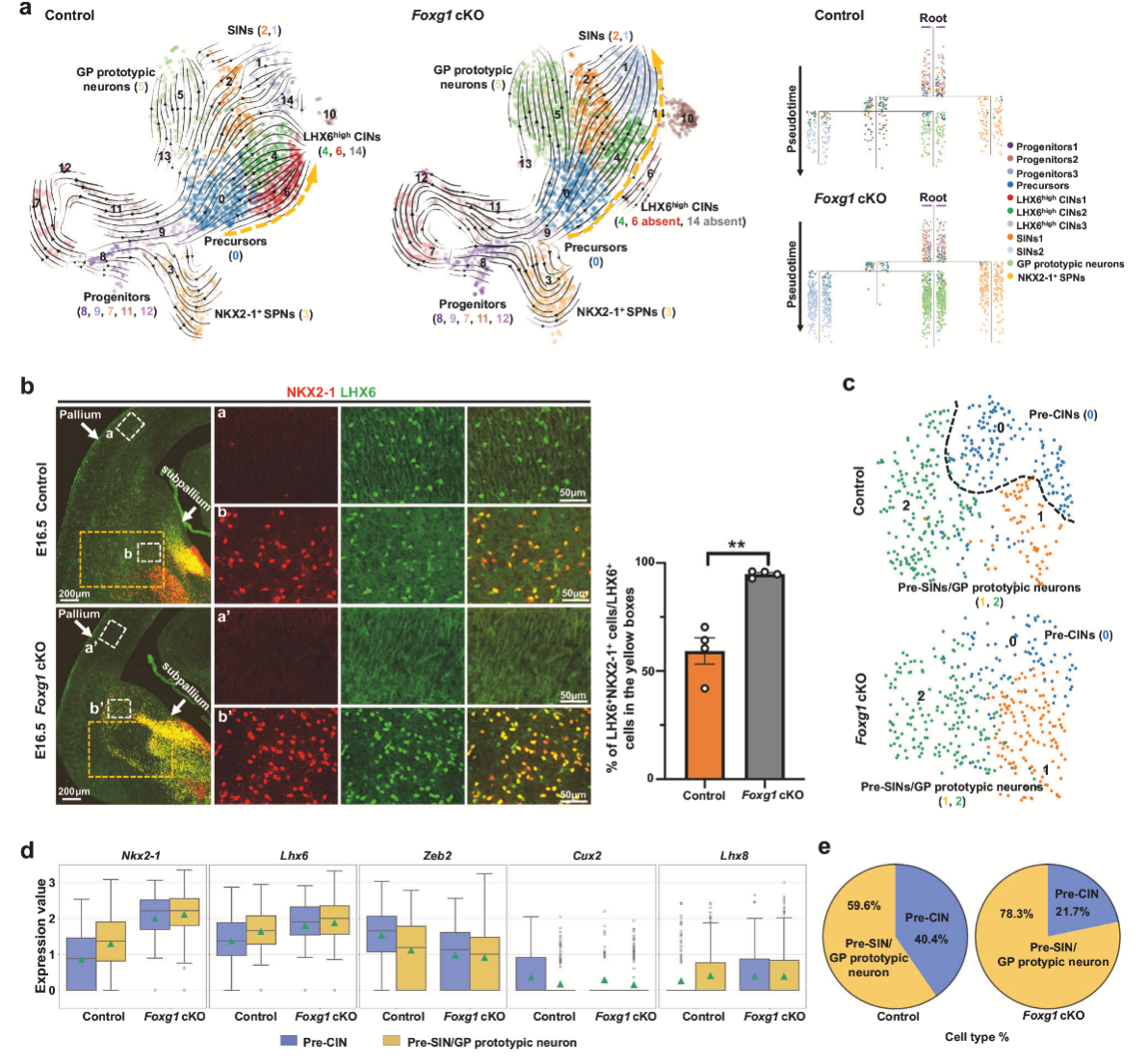
圖2 LHX6high CIN 向皮質(zhì)下區(qū)的轉(zhuǎn)化開始于 Foxg1cKO MGE 譜系的前體階段
3. FOXG1驅(qū)動的轉(zhuǎn)錄程序指導(dǎo) MGE 譜系中 LHX6high CIN與SIN/GP原型神經(jīng)元的命運選擇
為進(jìn)一步探究Foxg1調(diào)控MGE譜系神經(jīng)元發(fā)育的分子機(jī)制,通過RNA 速率分析,作者繪制了對照組MGE譜系神經(jīng)元亞型的完整發(fā)育軌跡,并鑒定出各軌跡中表達(dá)變化最顯著的500個基因。研究發(fā)現(xiàn),LHX6high CINs軌跡中Nkx2-1和Lhx8表達(dá)下調(diào),而Cux2、Tcf4等皮質(zhì)命運相關(guān)基因上調(diào);SINs和GP神經(jīng)元軌跡則呈現(xiàn)相反模式,其中Dach1和Pbx3分別特異性上調(diào)(圖3b)。
作者采用非負(fù)矩陣分解(NMF)方法,從對照組MGE譜系中定義了8個基因模塊,鑒定出56個調(diào)控神經(jīng)元命運的關(guān)鍵轉(zhuǎn)錄因子(如Tcf4、Dlx2、Zeb2),其中86%受FOXG1直接或間接調(diào)控(圖3d)。為驗證這些發(fā)現(xiàn),作者選擇關(guān)鍵模塊中的Sox4進(jìn)行功能驗證,發(fā)現(xiàn)在E12.5過表達(dá)Sox4能顯著增加E16.5時皮質(zhì)LHX6+ CINs的比例,這與Foxg1 cKO中Sox4下調(diào)導(dǎo)致CINs減少的表型相反,證實了Sox4促進(jìn)皮質(zhì)命運的作用(圖3e)。
這些結(jié)果共同表明,F(xiàn)oxg1通過直接調(diào)控核心轉(zhuǎn)錄因子網(wǎng)絡(luò)(如Sox4、Tcf4等),動態(tài)平衡皮質(zhì)與腹側(cè)命運相關(guān)基因的表達(dá),從而精確指導(dǎo)MGE譜系神經(jīng)元的分化選擇。
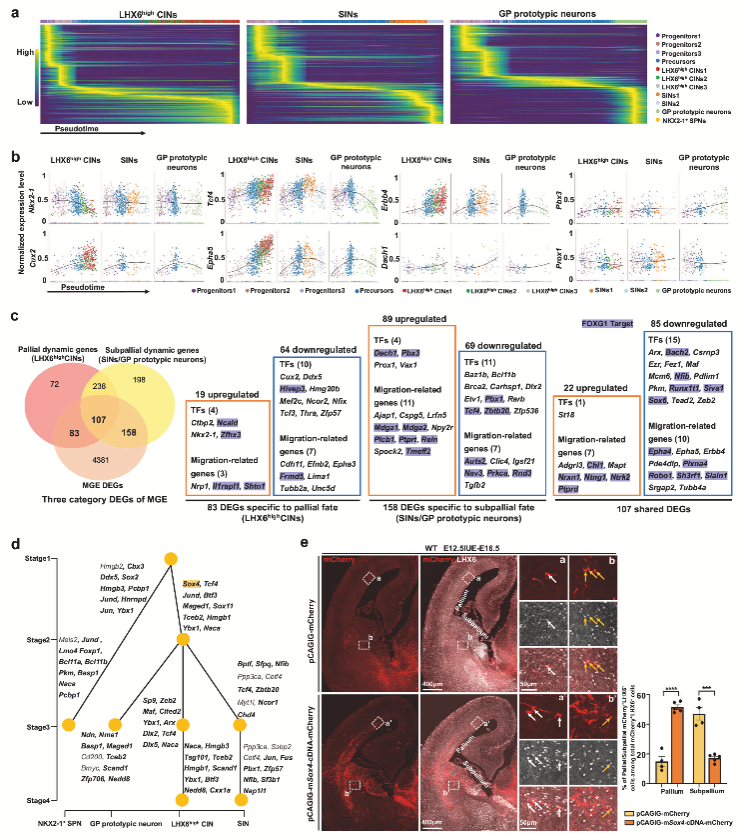
圖3 Foxg1對MGE譜系神經(jīng)元發(fā)育的影響
4. 在dLGE/CGE譜系中,ID3high OBIN 向GP Arkypallidal 神經(jīng)元轉(zhuǎn)化,SP8high OBIN/PROX1high CIN 向杏仁核神經(jīng)元轉(zhuǎn)化
為了深入解析Foxg1 cKO對dLGE/CGE譜系神經(jīng)元發(fā)育的調(diào)控機(jī)制,作者通過發(fā)育軌跡分析,發(fā)現(xiàn)dLGE/CGE細(xì)胞存在兩個獨立的發(fā)育譜系:FOXP2highSP8low譜系(產(chǎn)生ID3high OBINs和GP Arkypallidal神經(jīng)元)和FOXP2lowSP8high譜系(產(chǎn)生SP8high OBINs、PROX1high CINs和杏仁核神經(jīng)元)。免疫染色證實SP8與FOXP2在dLGE/CGE細(xì)胞中呈現(xiàn)互補表達(dá)模式(圖4a)。在FOXP2highSP8low譜系中,ID3high OBINs的發(fā)育軌跡明顯向GP Arkypallidal神經(jīng)元偏移,其發(fā)育分支縮短而GP神經(jīng)元分支延長(圖4c,d);在FOXP2lowSP8high譜系中,OBINs/CINs分支縮短而杏仁核神經(jīng)元分支延長,表明SP8high OBINs和PROX1high CINs轉(zhuǎn)向杏仁核神經(jīng)元命運(圖4c,d)。這些發(fā)現(xiàn)表明FOXG1和下游轉(zhuǎn)錄程序指導(dǎo)dLGE/CGE譜系中的命運選擇。
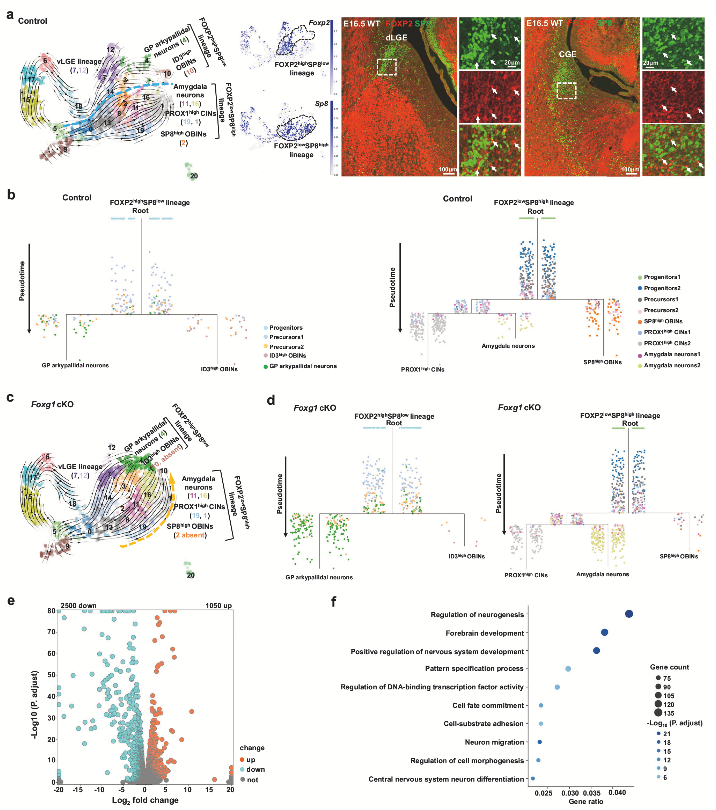
圖4 Foxg1對dLGE/CGE譜系神經(jīng)元發(fā)育的影響
5. FOXP2highSP8low和FOXP2lowSP8high dLGE/CGE 譜系中指定每種神經(jīng)元類型的獨特轉(zhuǎn)錄程序
隨后作者進(jìn)一步揭示了FOXP2highSP8low和FOXP2lowSP8high dLGE/CGE 譜系中的發(fā)育機(jī)制。分別鑒定各神經(jīng)元亞型發(fā)育過程中表達(dá)變化最顯著的500個基因(圖5a,b),在FOXP2highSP8low譜系中,ID3high OBINs和GP Arkypallidal神經(jīng)元在命運決定關(guān)鍵期表現(xiàn)出顯著差異:ID3high OBINs特異性上調(diào)Id2、Hivep3等轉(zhuǎn)錄因子和Tcf4、Erbb4等遷移相關(guān)基因(圖5a);而GP Arkypallidal神經(jīng)元則選擇性上調(diào)Tshz2并維持Prox1表達(dá)。FOXP2lowSP8high譜系中,SP8high OBINs、PROX1high CINs和杏仁核神經(jīng)元在發(fā)育中后期呈現(xiàn)明顯差異:SP8high OBINs激活Tshz1;PROX1high CINs中Nfix、Htr3a等皮質(zhì)中間神經(jīng)元特征基因表達(dá)上調(diào);杏仁核神經(jīng)元則特異性表達(dá)Wls等標(biāo)記物(圖5b)。
通過整合差異基因和CUT&Tag數(shù)據(jù),作者在FOXP2highSP8low譜系中鑒定出343個關(guān)鍵差異表達(dá)基因,其中34%為FOXG1直接靶標(biāo)(圖5c)。這些基因可分為三類:111個ID3high OBINs特異性基因(如下調(diào)的Tcf4、Epha4)、113個GP Arkypallidal神經(jīng)元特異性基因(如上調(diào)的Meis2、Tshz2)以及119個共有基因。NMF分析進(jìn)一步揭示34個核心轉(zhuǎn)錄因子構(gòu)成調(diào)控網(wǎng)絡(luò),其中88%受FOXG1調(diào)控(圖5e)。Meis2作為FOXG1直接靶標(biāo),其上調(diào)與腹側(cè)GP神經(jīng)元增加顯著相關(guān)。在FOXP2lowSP8high譜系中,研究鑒定出調(diào)控SP8high OBINs/PROX1high CINs與杏仁核神經(jīng)元命運分化的46個關(guān)鍵轉(zhuǎn)錄因子(圖5f),包括Etv1、Nr2f2等已知因子及St18、Hmgb1等新發(fā)現(xiàn)調(diào)控因子,其中85%(39個)受FOXG1調(diào)控。功能驗證實驗顯示,在E12.5過表達(dá)Hmgb1能顯著增加E16.5時皮質(zhì)PROX1+ CINs的比例(圖5g),與Foxg1 cKO中Hmgb1下調(diào)導(dǎo)致CINs減少結(jié)果一致。
這些發(fā)現(xiàn)共同表明,F(xiàn)oxg1通過差異調(diào)控兩個譜系的轉(zhuǎn)錄程序:在FOXP2high譜系中平衡Id2/Hivep3與Meis2/Tshz2的表達(dá)決定OBINs/GP神經(jīng)元命運;在FOXP2low譜系中通過Hmgb1等因子調(diào)控皮質(zhì)與腹側(cè)神經(jīng)元的分化選擇。
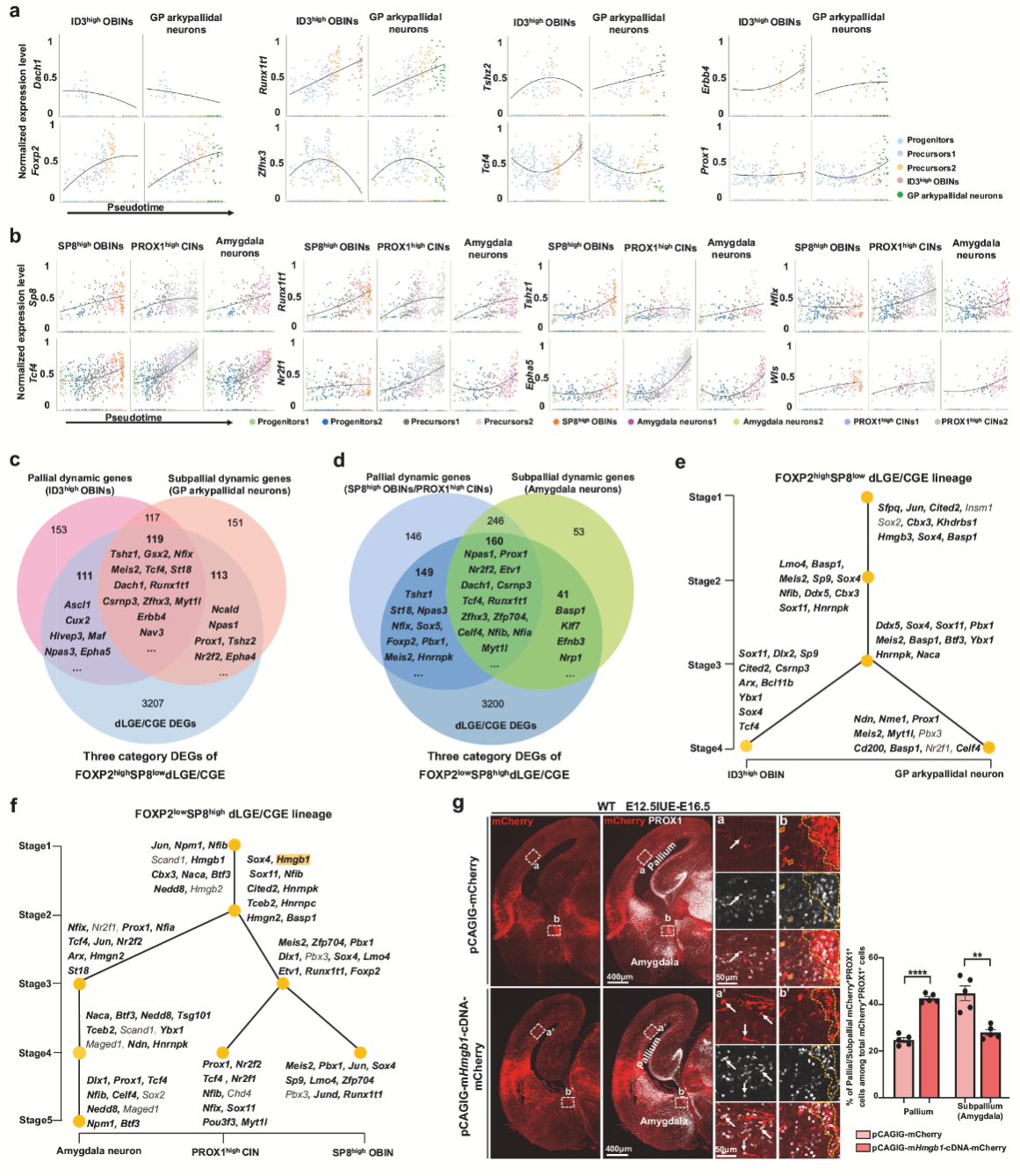
圖5 FOXP2highSP8low和FOXP2lowSP8high dLGE/CGE 譜系的轉(zhuǎn)錄組特征
6. Foxg1調(diào)控皮質(zhì)/皮質(zhì)下區(qū)GABA能神經(jīng)元發(fā)育的統(tǒng)一機(jī)制
通過整合MGE和dLGE/CGE譜系的差異表達(dá)分析,作者鑒定出40個共同調(diào)控因子(包括Tcf4、Prox1等19個FOXG1直接靶標(biāo)),其中32個與自閉癥、精神分裂癥等神經(jīng)精神疾病相關(guān)(圖6a,b)。作者選擇自閉癥相關(guān)基因Tcf4進(jìn)行深入解析:熒光素酶實驗證實FOXG1直接激活Tcf4啟動子(圖7b);在Foxg1 cKO中恢復(fù)Tcf4表達(dá)能顯著增加皮質(zhì)PROX1+ CINs的數(shù)量(圖7c)。這些結(jié)果不僅證實Tcf4是FOXG1調(diào)控神經(jīng)元命運的關(guān)鍵效應(yīng)分子,更揭示了神經(jīng)精神疾病風(fēng)險基因如何通過干擾GABA能神經(jīng)元命運決定參與病理過程。
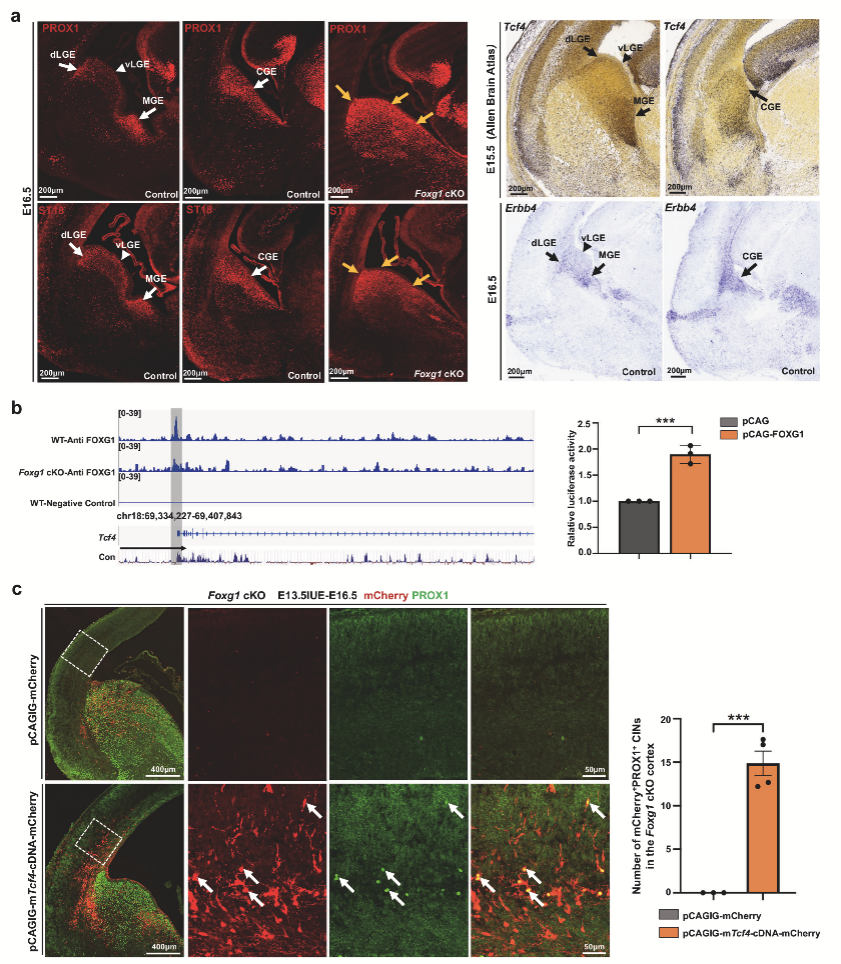
圖6 Foxg1 cKO CGE中恢復(fù)Tcf 4挽救了皮質(zhì)PROXl+CIN命運
參考文獻(xiàn):
[1] Sun L, Xiong F, Huang F, Dong S, Zhu P, Jiang P, Zhang B, Zhang X, Sun J, Peng H and Zhao C 2025 Transcriptomic insights into fate choice of pallial versus subpallial GABAergic neurons Nat Commun 16 5032