原代肝細胞是一類從動物肝臟獲得后立即培養的肝細胞。由成人原代肝細胞(adult primary human hepatocyte, PHH) 分化培養得到的類器官因保留有體內生理環境、肝細胞的完整形態和生理功能,往往被視作為體外代謝藥物實驗與體外肝細胞培養模型的“金標準”,有助于研究藥物代謝和毒性、病毒感染和遺傳疾病等。
然而,體外培養的PHH往往只能保留幾天的代謝活性,隨后將會去分化并凋亡。為此,研究人員探索了各類可能的替代方法。例如,使用層膠原凝膠或Matrigel包裹原代肝細胞的2D“三明治培養”;或與肝臟非實質細胞共培養,但效果皆不盡如人意。而使用其他來源的肝細胞替代,如癌細胞系來源,雖會保有增殖能力、但缺乏正常肝細胞關鍵特征。
2013年,荷蘭研究團隊在培養體系中加入肝細胞生長因子 (HGF)、表皮生長因子 (EGF)、成纖維細胞生長因子 (FGF) 等組分后,成功建立了老鼠來源的肝臟類器官體系。可惜多種藥物在動物模型上和臨床表現存在較大出入。而來自于人多能干細胞或胎兒肝細胞 (human fetal hepatocytes, FH) 的模型也被發現會產生巨噬細胞招募因子與抗纖維化M2巨噬細胞極性因子高度表達等與PHH有顯著區別的特性。
2024年5月13日,來自荷蘭烏特勒支大學,被譽為“類器官之父”的Hans Clevers教授,帶領團隊在期刊Nature Communications上發表了題為“Mapping of mitogen and metabolic sensitivity in organoids defines requirements for human hepatocyte growth”的研究論文。

通過追蹤人類來源成人肝細胞培養中的時間轉錄組和表型變化,并將其與胎兒來源肝細胞進行比較,研究團隊首次發現基于成人肝細胞的類器官培養中存在增殖啟動和脂質代謝抑制的缺失,并證明在補充IL6+FXRa等生長相關因子后,具有成年肝細胞特征的類器官可在體外培養長達四個月,為基于PHH的類器官體外培養提供了新的線索。
細胞增殖基因與脂肪代謝基因表達成動態相反趨勢
研究團隊選取來自兩個供體的FH,在播種后三天內成功觀察到小型類器官組織的出現,并經熒光鑒定確認甲胎蛋白和白蛋白等肝細胞特異性蛋白的存在,同時發現膽管細胞的缺失。
其后根據時間分辨轉錄組學結果,研究人員將相關基因分為7簇。其中基因簇2與基因表達和DNA復制有關,其表達量會在早期時間點迅速增加,準備傳代時降低,而在傳代后再次升高,反映了動態的增殖反應;而與脂質代謝過程相關的基因簇7中則呈現了截然相反的態勢,即在早期時間點表達迅速減少,準備傳代時增加,傳代后再次下降。
為了進一步探究單純肝細胞中的動態變化,研究人員通過獲取單細胞FH并再次進行了類器官再生的早期轉錄組觀測,結果與此前保持一致。另外,形態學上,早期單個FH只表現脂肪積累,并不進行復制增殖;48小時內,FH完成第一輪分裂,脂質表型在此時同步消失;培養三天后,無脂質的多細胞FH類器官最終形成。該形態學變化反映脂質代謝信號確乎與生長相關信號存在有緊密關聯。
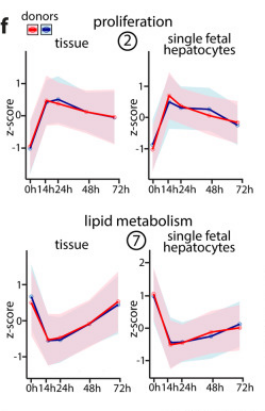
圖1:FH類器官生長過程中,組織簇2和7的RNA-seq結果
使用生長信號及細胞因子優化FH類器官培養基
對于基因簇2而言,Wnt被認為深度參與細胞的增殖與分化。當移除Wnt信號通路激活劑RSPO1后,類器官生長無顯著影響;而移除另一激活劑CHIR后,FH類器官則會迅速凋亡。此外,在添加cAMP激活劑forskolin (FSK) 和糖皮質激素類似物地塞米松 (Dex) 后誘導分化后,膽管細胞類器官生長被激活,肝細胞類器官則表現出多角形形態、體積增大等成熟特質。
除此以外,細胞因子也可顯著促進FH類器官生長,具體效果則是NRG1>IL6>IL11。
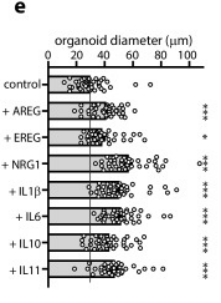
圖2:添加不同生長因子的FHs在從單個FHs出芽7天后的直徑對比
PHH類器官培養中的停滯
完成FH相關機制的探索之后,研究人員很快在PHH上觀察到了不一樣的轉錄組圖譜。PHH會在長出小型類器官后停止增長,且在傳代后也無法恢復增長;與此同時,基因簇2和7的表達水平在傳代后保持平穩,而非升高(基因簇2)或降低(基因簇7)到早期水平,指示增殖與脂質代謝無法重啟。經典衰老標志物CDKN1A (p21)的發現,也證實體外培養的PHH確已進入生長停滯狀態。
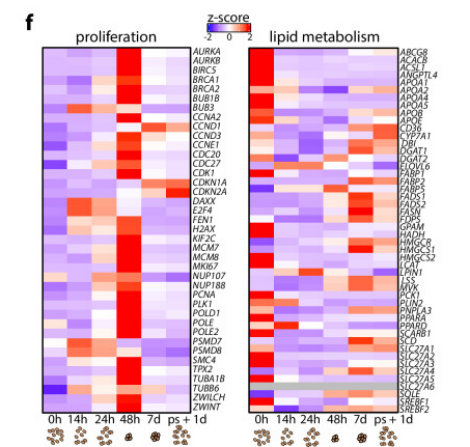
圖3:PHH類器官生長過程中基因簇2和7表達熱圖
生長因子IL6及脂質代謝因子FXRa誘導PHH體外發育
通過探索FH類器官培養中得到的思路,研究人員試圖添加類似的生長因子進行誘導。在FHs上表現卓越的生長因子中,只有白介素6 (IL6) 對PHH類器官表現出了顯著的激活效應,觀測到明顯的類器官直徑增加。IL6由巨噬細胞、T細胞、B細胞等多細胞產生,深度參與并調節多種細胞的生長與分化、免疫應答與炎癥反應等。此前已經發現血液IL6水平會于部分肝的切除術后迅速增加,并顯著影響肝切除后再生的效果與速度。且有文章報道,IL6可以將肝細胞轉化為保持分化為肝細胞能力的誘導性肝祖細胞 (iHPCs),促進小鼠原代肝細胞在體外的長期擴展,支持建立單個肝細胞衍生的iHPC克隆等。因此,本研究將IL6的促進生長效果推演到了輔助PHH類器官體外培養的有效條件,論證其可作為體外肝細胞長期培養的支持組分之一。
不過,當只添加IL6時,PHH類器官仍表現出與生長同步的脂質積累,而細胞分化似乎未被誘導成功。研究團隊隨后加入了法尼醇X受體 (farnesoid X receptor, FXR) 激動劑。FXR是一類膽汁酸受體,其中a1/2亞型主要在肝臟和腎上腺中表達并調控脂質代謝。此前有動物實驗表明,激活FXR可顯著降低游離脂肪酸、甘油三酯和總膽固醇等積累,并呈現質量效應。而使用非甾體FXR激動劑Cilofexor則會降低人類肝脂肪變性和血清膽汁酸水平。本文使用FXRa抑制脂質積累,變相進一步促進了PHH類器官發育。
此外,為了消除潛在的衰老標志物p21,研究團隊還添加了經典的類器官培養細胞因子Noggin。Noggin可抑制BMP信號,以保持維持肝前體細胞未分化狀態,催動其向成熟肝細胞和膽管細胞分化。
最終的培養基方案被證實可持續擴增PHH類器官至少四個月。
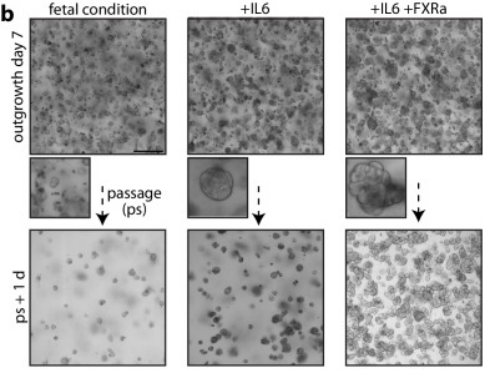
圖4:PHHs的培養基優化
總結與展望
本研究中,研究團隊追蹤FH類器官培養中的時間轉錄組和表型變化,發現了增殖啟動和脂質代謝抑制的同步性。并據此對PHH類器官的培養條件進行了模擬優化,通過IL6促進增殖啟動、FXRa抑制脂質代謝、Noggin抑制衰老輔助分化,成功達成了PHH類器官體外四個月培養的階段性成果,單細胞RNA測序也證實了其成年肝細胞身份。這一結果不僅揭示了決定肝細胞從發育到成年增殖的有絲分裂原需求和代謝差異,也為進一步探索體外PHH類器官的構建提供了重要依據。
不過作者也在文中特別提到,該研究中所使用的肝細胞來源人數有限,且捐贈者整體年齡偏低,未來需要在更多捐獻者中進行個性化多樣化測試,以確保藥物利用的普適性。
參考資料
1. Qiu L, Kong B, Kong T, Wang H. Recent advances in liver-on-chips: Design, fabrication, and applications. Smart Med. 2023 Feb 12;2(1):e20220010. doi: 10.1002/SMMD.20220010. PMID: 39188562; PMCID: PMC11235950.
2. Antarianto RD, Mahmood A, Giselvania A, Asri Dewi AAP, Gustinanda J, Pawitan JA. Inventing Engineered Organoids for end-stage liver failure patients. J Mol Histol. 2022 Aug;53(4):611-621. doi: 10.1007/s10735-022-10085-7. Epub 2022 Jul 27. PMID: 35882727; PMCID: PMC9374785.
3. Kaur I, Vasudevan A, Rawal P, Tripathi DM, Ramakrishna S, Kaur S, Sarin SK. Primary Hepatocyte Isolation and Cultures: Technical Aspects, Challenges and Advancements. Bioengineering (Basel). 2023 Jan 18;10(2):131. doi: 10.3390/bioengineering10020131. PMID: 36829625; PMCID: PMC9952008.
4. Nuciforo S, Heim MH. Organoids to model liver disease. JHEP Rep. 2020 Oct 22;3(1):100198. doi: 10.1016/j.jhepr.2020.100198. PMID: 33241206; PMCID: PMC7672322.
5. Tadokoro T, Murata S, Kato M, Ueno Y, Tsuchida T, Okumura A, Kuse Y, Konno T, Uchida Y, Yamakawa Y, Zushi M, Yajima M, Kobayashi T, Hasegawa S, Kawakatsu-Hatada Y, Hayashi Y, Osakabe S, Maeda T, Kimura K, Mori A, Tanaka M, Kamishibahara Y, Matsuo M, Nie YZ, Okamoto S, Oba T, Tanimizu N, Taniguchi H. Human iPSC-liver organoid transplantation reduces fibrosis through immunomodulation. Sci Transl Med. 2024 Jul 24;16(757):eadg0338. doi: 10.1126/scitranslmed.adg0338. Epub 2024 Jul 24. PMID: 39047116.
6. Shafritz DA, Ebrahimkhani MR, Oertel M. Therapeutic Cell Repopulation of the Liver: From Fetal Rat Cells to Synthetic Human Tissues. Cells. 2023 Feb 6;12(4):529. doi: 10.3390/cells12040529. PMID: 36831196; PMCID: PMC9954009.
7. Cressman DE, Greenbaum LE, DeAngelis RA, Ciliberto G, Furth EE, Poli V, Taub R. Liver failure and defective hepatocyte regeneration in interleukin-6-deficient mice. Science. 1996 Nov 22;274(5291):1379-83. doi: 10.1126/science.274.5291.1379. PMID: 8910279.
8. Guo R, Jiang M, Wang G, Li B, Jia X, Ai Y, Chen S, Tang P, Liu A, Yuan Q, Xie X. IL6 supports long-term expansion of hepatocytes in vitro. Nat Commun. 2022 Nov 29;13(1):7345. doi: 10.1038/s41467-022-35167-8. PMID: 36446858; PMCID: PMC9708838.
9. 陳立新,林創珍,郁冰清,等.奧貝膽酸治療非酒精性脂肪肝的療效和機制——膽汁酸藥物的利和弊[J].中國臨床新醫學,2021,14(8):756-761.
10. Hendriks D, Artegiani B, Margaritis T, Zoutendijk I, Chuva de Sousa Lopes S, Clevers H. Mapping of mitogen and metabolic sensitivity in organoids defines requirements for human hepatocyte growth. Nat Commun. 2024 May 13;15(1):4034. doi: 10.1038/s41467-024-48550-4. PMID: 38740814; PMCID: PMC11091073.
11. Yuan X, Wu J, Sun Z, Cen J, Shu Y, Wang C, Li H, Lin D, Zhang K, Wu B, Dhawan A, Zhang L, Hui L. Preclinical efficacy and safety of encapsulated proliferating human hepatocyte organoids in treating liver failure. Cell Stem Cell. 2024 Apr 4;31(4):484-498.e5. doi: 10.1016/j.stem.2024.02.005. Epub 2024 Mar 7. PMID: 38458193.
12. 劉文明,鄢和新.淺析類器官與肝臟再生研究進展[J].肝臟,2023,28(06):637-638.