植物激素是調節植物生長、發育和環境應激反應等關鍵方面的信號化合物。干旱、鹽堿、高溫、寒冷和洪澇等非生物脅迫對植物的生長和存活有著深遠的影響。對這種脅迫的適應和耐受涉及到復雜的感知、信號和脅迫反應機制。
2022年5月6日,Nature reviews molecular cell biology(IF=94.4 2021年)雜志在線發表了美國科學院院士、中科院特聘研究員、美國加州大學圣地亞哥分校Julian Schroeder教授為通訊作者的題為“Plant hormone regulation of abiotic stress responses”的綜述文章。

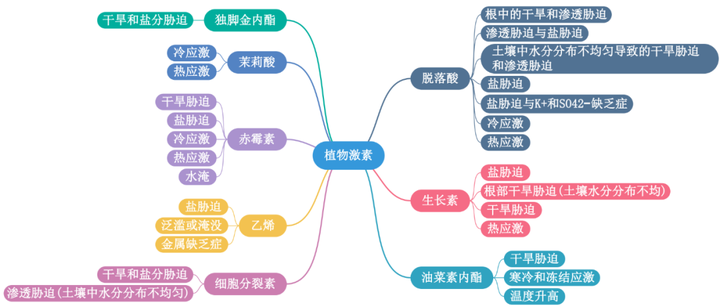
植物在不斷變化的環境條件發展出復雜的機制來感知和響應外部壓力。其中,非生物脅迫以各種形式出現,為了保護植物免受非生物脅迫負面影響,我們需要在分子水平上了解植物脅迫反應的機制。植物種中9種常見的植物激素:脫落酸(ABA)、生長素、油菜素內酯、細胞分裂素、乙烯、赤霉素、茉莉酸、水楊酸和獨角金內酯,是眾所周知的植物生長調節劑,本文首先列舉了這些植物激素在非生物脅迫反應中最顯著的作用。
百趣提供全面包含九大類植物激素的植物激素高通量檢測方案,采用定量金標準-同位素內標法絕對定量,詳細內容可點擊鏈接查看:技術介紹|植物激素高通量靶標定量
隨后作者主要圍繞非生物脅迫(包括干旱、滲透脅迫和水澇)從植物如何感受脅迫、介導植物非生物脅迫反應激素依賴的基因調控機制、脅迫相關激素反應對種子萌發和開花時間的影響、ABA和生長素如何協調脅迫下的根生長、乙烯和赤霉素介導的植物對淹水反應的調節、ABA和非生物脅迫感應機制調節氣孔開度等方面全面描述了植物激素和其他信號化合物如何調節植物對非生物脅迫的反應的研究現狀。
滲透脅迫傳感與信號轉導
包括植物從外部土壤中吸收水分以及植物內部水分的轉運均是由水勢梯度決定的,例如水澇等低滲脅迫下會導致細胞腫脹,而干旱鹽堿等高滲脅迫下則會導致植物萎蔫。植物已經進化出包括通過依賴ABA和不依賴ABA的途徑調節細胞滲透濃度、氣孔運動和植物發育等滲透脅迫適應機制。
1. 滲透和鹽脅迫傳感機制
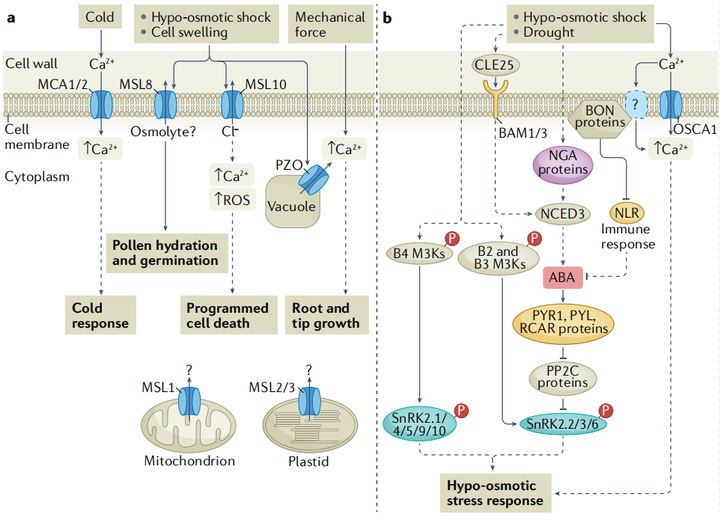
植物之所以能夠感知膨壓變化、細胞內溶質濃度的輕微變化以及滲透脅迫對細胞結構的機械作用,是因為鈣信號在滲透脅迫中起到了關鍵作用,植物細胞內游離鈣濃度([Ca2+]cyt)在受到滲透脅迫后的幾秒鐘內會迅速且瞬時地升高。早已有研究表明了機械敏感離子通道在滲透脅迫傳感機制中的作用(圖2a)。比如類似小導電的機械敏感通道(mechanosensitive channel of small conductance-like MSL)是細胞器低滲、花粉水合和萌發、根中的觸摸反應和細胞膨大過程中由膜張力激活的非選擇性離子通道;MID1-補充活動型鈣滲透通道(mid1-complementing activity MCA)會被膜張力激活,進而介導根的低滲透休克和觸覺感知;以及機械敏感離子通道PIEZO1(PZO1)是根尖機械轉導所必需 ,可以介導擬南芥花粉管尖端的液泡管狀形成;OSCA1(reduced hyperosmolality-induced[Ca2+]cyt increase 1)是一種潛在的滲透壓感受器,最典型的特征是在高滲透壓下激活Ca2+陽離子通道(圖2b)。最近的研究表明鈣響應型磷脂結合蛋白(Ca2+-responsive phospholipid-binding BONZAI BON)通過正向調節滲透脅迫誘導的[Ca2+]cyt增加、ABA積累和基因表達來介導高滲透脅迫耐受性,這些膜相關的鈣響應性BON蛋白可能通過與質膜鈣轉運蛋白一起調節初始的[Ca2+]cyt升高而參與滲透感知和信號轉導。
植物感知鹽度并誘導快速而瞬時的[Ca2+]cyt升高,進而通過鹽過度敏感的途徑觸發耐鹽反應(圖2c)。鹽脅迫下,受體樣激酶Feronia能夠感知鹽度引起的細胞壁缺陷,并引發細胞特異性的[Ca2+]cyt細胞信號以維持細胞壁的完整性。Feronia還可能通過與ABI2(protein phosphatase 2c(PP2C)ABA-insensitive 2)的相互作用來干擾ABA信號。
2. 滲透脅迫誘導ABA的生物合成
內源性ABA濃度在植物缺水后約2.5-6小時內會逐漸升高。由NCED3基因編碼9-cis-epoxycatoteinoid dioxygenase會催化ABA從頭合成的限速步驟,同樣的,NCED3在葉綠體中的翻譯后處理也有被報道能夠調節ABA的積累。作為對根缺水的響應,水勢信號參與了從根到冠的快速缺水信號,從而觸發擬南芥葉片和氣孔關閉中ABA的生物合成。在轉錄水平上,擬南芥NAC轉錄因子ATAF1被認為調節NCED3的表達,以促進ABA的積累(圖2b)。
非生物脅迫下的基因調控
基因表達的變化介導了植物激素的許多作用。 早期的基因組技術表明,與非生物脅迫相關的ABA水平升高會改變數千個基因的mRNA水平,連同許多經典的ABA不敏感突變被定位到轉錄調節因子的發現,表明基因調控在非生物脅迫抗性中的重要作用。
1.ABA介導的轉錄調控與激素串擾
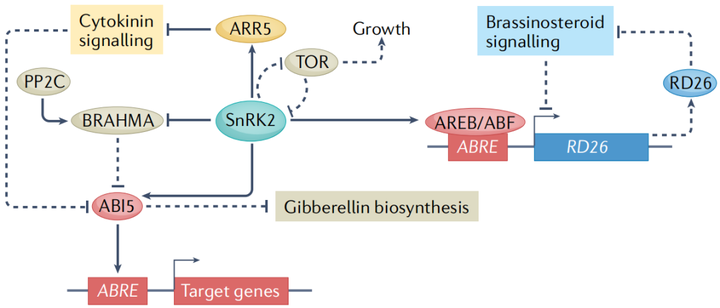
早期的研究發現,在干旱誘導基因的啟動子中有一種保守的順式作用調控元件,稱為ABA反應元件(ABRE)。ABREs被堿性亮氨酸拉鏈型轉錄因子識別 (圖3)。在ABA信號轉導過程中,依賴于ABA的SnRK2型蛋白激酶直接磷酸化并激活AREBs/ABF(ABRE-binding proteins/ABRE-binding factors)和ABI5。一項利用染色質免疫沉淀和測序的開創性研究,在ABA反應期間分析了21個轉錄因子的全基因組結合位點,發現許多結合事件是動態的,并且多個轉錄因子可以針對同一基因。重要的是,ABA誘導的某些轉錄因子的結合與相鄰ABRE位點的存在呈正相關,這表明一些轉錄因子可能與AREBs/ABF協同作用。
在沒有非生物脅迫的情況下,不同的激素途徑相互作用以控制植物生命活動的許多方面。比如在干旱脅迫方面,ABA誘導的干燥反應轉錄因子26(responsive to desiccation 26 RD26)抑制油菜素內酯誘導的基因的一個子集,同時,油菜素內酯激活的轉錄因子則會抑制RD26的表達,表明ABA信號和油菜素內酯信號之間的拮抗串擾有助于干旱脅迫反應(圖3);細胞分裂素與ABA也同樣存在串擾現象,SnRK2介導的A型擬南芥反應調節因子5(type-A arabidopsis response regulator 5 ARR5)的磷酸化是細胞分裂素信號傳導的負調節因子,可促進其蛋白質穩定性,從而在干旱脅迫期間下調細胞分裂素反應。
2. 轉錄后非生物脅迫反應
轉錄后過程將基因調控的范圍進一步擴展到轉錄控制之外,最近的研究揭示了這些機制在塑造ABA反應中的作用。最近發現一種ABA超敏突變體含有編碼ABA超敏1(ABA hypersensitive 1 ABH1)mRNA帽結合蛋白的基因,使得mRNA處理和ABA信號之間建立了早期的聯系,表明選擇性的mRNA剪接會受到非生物脅迫的調節,并調節ABA的反應。根據近期的研究報道,mRNA衰變已成為導致非生物應激反應的額外機制。mRNA分子的降解是由mRNA脫帽介導的。在滲透脅迫期間,ABA非依賴的SnRK2蛋白激酶會使解帽激活物VARICOSE(VCS)磷酸化,從而導致某些轉錄物的不穩定。
非生物脅迫下的生長調控
植物激素調節植物生長發育的許多方面。最近的發現已經開始闡明它們如何控制植物生長和發育的不同策略,以響應非生物脅迫。
1.TOR與非生物脅迫的相互作用
雷帕霉素靶點(target of rapamycin TOR)是植物中一種重要的發育和代謝調節因子。ABA途徑和TOR途徑之間的相互調節來協調植物的生長和非生物脅迫反應。在非生物脅迫條件下,TOR使PYL/rcar ABA受體磷酸化,抑制ABA信號轉導,促進生長,而ABA激活的SnRK2型蛋白激酶磷酸化TOR 1B調節相關蛋白(regulatory- associated protein of tor 1B RAPTOR1B),抑制TOR激酶活性進而抑制生長(圖3)。
2.赤霉素、ABA和發芽的決定
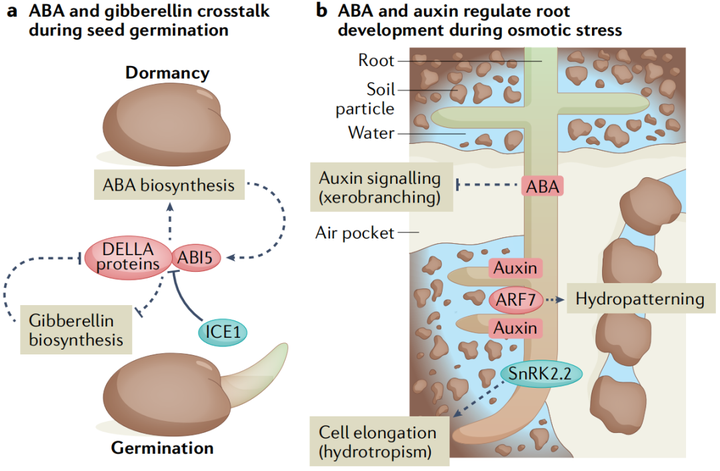
種子萌發的調節通過協調胚胎發育和出苗與環境條件來促進幼苗存活。兩種相互競爭的激素信號通路赤霉素和ABA的平衡決定了發芽的與否。在種子成熟期間,轉錄因子網絡包括ABA調節的轉錄因子ABI3、ABI4和ABI5,會誘導種子保持干燥和 ABA生物合成所需的基因,并抑制赤霉素生物合成基因。而植物觸發種子打破種子休眠的諸如冷和光等環境信號,是通過將平衡轉向赤霉素來實現的(圖4A)。ABA和赤霉素之間的這種拮抗關系出現在它們各自途徑的多個節點上,DELLA蛋白是植物特有的GRAS(GIBBERELLIN-INSENSITIVE,
REPRESSOR of ga1-3, SCARECROW)家族的成員。DELLA蛋白會抑制赤霉素反應,而赤霉素信號通過觸發DELLA蛋白的蛋白酶體降解部分使它們失活。DELLA蛋白與ABI3和ABI5相互作用,這些蛋白復合體共同刺激SOMNUS的轉錄,SOMNUS是一種關鍵的休眠促進因子,能夠激活ABA生物合成基因并抑制赤霉素生物合成基因。有趣的是,DELLA-ABI5復合體的作用被CBF表達的誘導物1(inducer of cbf expression 1 ICE1)抑制。ICE1的結合阻斷了ABI5的DNA結合活性,這種相互作用受到赤霉素處理的刺激,可能是由于DELLA蛋白的降解,通過該機制,預先暴露于低溫可能會促進發芽。
除了上述機制之外,ABI5的表達調控似乎是種子萌發過程中多種環境信號的主要調節點,光信號成分(elongated hypcotyl 5 HY5)響應光直接激活ABI5轉錄;DELLA蛋白RGL2通過增強ABI5的轉錄進一步促進ABA信號傳導;根據最新報道,高鹽度可能通過兩種轉錄因子AGL21和RSM1在暴露于NaCl期間增強ABI5的表達進而抑制種子萌發。
3. 生長素、ABA與脅迫下的根系生長
雖然高濃度的外源ABA抑制了根的生長,但較低濃度的ABA會刺激初級根的生長。土壤中的水分分布是不均勻的,植物通過向水性,在一定程度上解決了這種情況。ABA缺乏突變體的向水性受到損害,對于向水性,蛋白激酶SnRK2.2是通過促進根伸長區的皮質細胞伸長使植物根系的差異生長 (圖4B)。而低濃度的ABA通過AHA2(AUTOINHIBITEDH+-ATPASE 2)減弱pp2c對質外體H+外流的抑制,從而刺激初生根的生長。在高鹽度環境中,側根進入長時間的生長停滯,這需要內皮層ABA信號。根在遠離高鹽區域也表現出優先生長這一現象被稱為--鹽生(halotropism)。鹽處理誘導生長素轉運蛋白PIN2(PIN-FORMED 2)的內化,當根遇到縱向鹽度梯度時,生長素在離鹽源最遠的根一側積累,然后導致根彎曲。有趣的是,親水性似乎并不是通過生長素的重新分配來起作用的,這表明親鹽性是一個與眾不同的過程。
ABA還調節水分脅迫期間的根組織模式。內胚層ABA信號通過誘導microRNAs,miR165和miR166的表達來刺激木質部分化,miR165和miR166是維管發育的兩個關鍵調節因子。ABA在木質部細胞中也發揮作用,在那里它激活幾個促進木質部分化的維管相關NAC結構域(vascular-related NAC domain VND)轉錄因子的表達。
4. 赤霉素、ABA和乙烯調節非生物脅迫下的開花
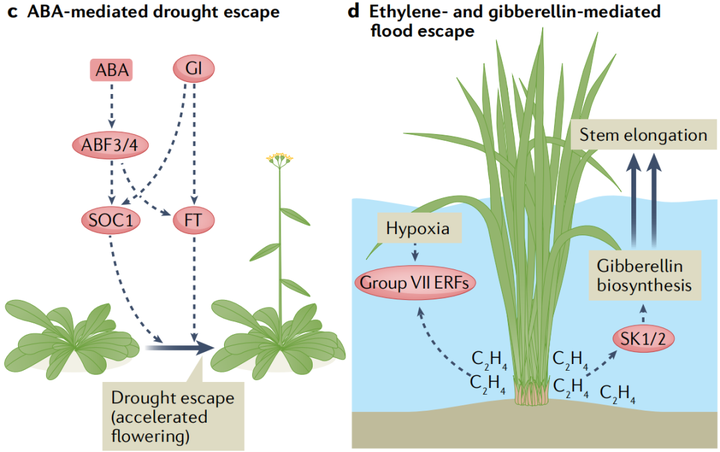
在長期干旱期間,許多植物會加速開花過程以在死亡前繁殖,這種反應被稱為干旱逃逸(drought escape)。雖然ABA在開花轉換過程中的確切作用目前尚不清楚。圍繞干旱加速開花的現象,新出現的證據表明ABA信號具有積極的作用。在長日照條件下,ABA生物合成突變體的開花時間延遲,而對ABA敏感的pp2c三重突變體118的開花時間提前。至關重要的是,干旱脅迫放大了這種延遲,表明植物需要ABA來促進干旱逃逸。這種ABA在干旱誘導的開花中的積極作用需要依賴于光周期的核心開花調節蛋白GI(GIGANTEA)(圖4c)。
與干旱相反,鹽脅迫會導致擬南芥中依賴乙烯的開花時間延遲。鹽脅迫通過誘導乙烯生物合成基因的表達導致乙烯積累。盡管潛在的機制尚不清楚,但乙烯干擾了赤霉素的信號傳遞,導致DELLA蛋白的積累,DELLA蛋白可以通過抑制開花刺激轉錄因子CONSTANS來推遲開花。此外,鹽脅迫還通過誘導GI的降解來抑制開花。
5. 乙烯和赤霉素介導的水澇脅迫響應
植物組織的淹沒阻礙了細胞對O2和CO2的獲取,這會嚴重擾植物的亂新陳代謝。此外,水下氣體擴散受限會導致乙烯在淹水的植物組織中積聚。長時間的淹水會導致低氧,這會激活一種保守的基因表達程序,支持植物在有限的氧氣中存活。一些適應洪水的物種表現出一種逃生策略,即水下的嫩芽和樹葉伸長到空中。對一種名為深水水稻(deepwater rice)的耐澇水稻品種的研究已經開始揭示乙烯和赤霉素信號是如何控制這種水下生長反應的。赤霉素通過促進節間生長來促進莖的伸長,這種關系在植物馴化過程中已經被利用。在深水水稻中,乙烯在淹沒的莖和葉組織中積累,這種升高的乙烯濃度誘導ERF編碼基因SNORKEL1和SNORKEL2的表達,這兩個主要數量性狀基因與深水水稻節間伸長有關。SNORKEL1和SNORKEL2可能通過誘導赤霉素的生物合成來刺激莖的伸長(圖4d)。最近,另一個與節間伸長相關的基因被定位到赤霉素生物合成的關鍵基因SEMIDWARF1(SD1)。與更常見的攜帶SD1零等位基因的半矮稈水稻品種不同,深水水稻植株誘導淹沒組織中SD1的表達。由此產生的赤霉素水平的增加與另一個稱為節間伸長加速器1(accelerator of internode elongation 1 ACE1)的基因一起作用,促進了中間分生組織中的細胞分裂。
氣孔運動的調節
葉表皮中保衛細胞形成的氣孔孔可以吸收二氧化碳進行光合作用,以換取水分。為了優化植物的水分利用效率,保衛細胞感知和響應包括光、二氧化碳和干旱在內的幾種非生物因素。
1.氣孔對干旱的響應
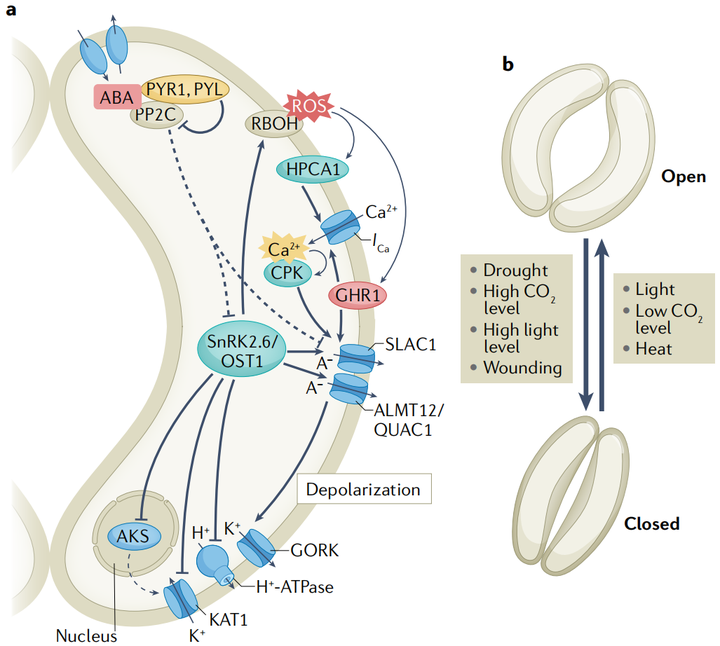
據報道,干旱脅迫會觸發維管組織和保衛細胞的ABA合成。保衛細胞中的ABA信號調節質膜離子通道,觸發負離子和K+的長期外流,導致保衛細胞縮小和氣孔關閉。保衛細胞的負離子釋放和隨后的質膜去極化是由慢型和快型陰離子通道介導的。擬南芥保衛細胞中一個主要的慢型陰離子通道由慢陰離子通道相關1(slow anion channel- associated 1 SLAC1)編碼, 陰離子通道觸發的去極化繼而通過電壓依賴K+通道GORK誘導K+流出。蛋白激酶SnRK2.6是保衛細胞中ABA信號傳導的主要正調節因子。SnRK2.6/OST1磷酸化并激活SLAC1和ALMT12 /QUAC1。A組pp2c蛋白作為負性ABA信號調節劑,不僅直接去磷酸化SnRK2.6/OST1 ,而且還使SLAC1失活(圖5a)。
2. 保衛細胞中的非生物信號整合
保衛細胞可以感知和整合幾種環境刺激(圖5b)。其中,光和CO2是調節氣孔開度的主要非生物刺激。藍光和紅光誘導氣孔開放機制,使光合作用最大化。光誘導的氣孔開放是由H+-ATPase酶激活和隨后通過保衛細胞膜上的電壓依賴的內向整流鉀(K+in)通道吸收K+所介導的。ABA通過抑制H+-ATPase酶和通道中的K+來抑制光誘導的氣孔開放。D組pp2c蛋白及其負調控因子小生長素上調RNAs(SAUR)也參與了擬南芥保衛細胞中H+-ATPase酶的調節。
ABA和其他激素之間的信號串擾有助于保衛細胞的非生物脅迫反應,據報道,過量的強光脅迫觸發了局部和全株系統的氣孔關閉,這可能是由NADPH氧化酶RBOHD與ABA、水楊酸和茉莉酸信號協同介導的。然而,黑暗和高二氧化碳濃度并不會導致擬南芥系統葉片的氣孔關閉。在高溫脅迫下,植物打開氣孔,通過蒸騰作用為葉片降溫。茉莉酸可以在高溫和其他例如強光和機械損傷脅迫的組合中微調氣孔。
研究小結
由于氣候變化,干旱、鹽分、高溫和洪澇等非生物脅迫對植物的挑戰越來越大。氣候變化和非生物脅迫也會加劇植物病害。植物生物學的最新進展為植物如何感知和響應非生物脅迫提供了至關重要的新見解。雖然將這些發現轉化為田間應用仍然具有挑戰性,但本文對個體激素調控的非生物脅迫反應的深入理解,有可能為選育更具抗性的作物提供關鍵的見解。闡明控制這些性狀的機制、基因和途徑可以為應用和轉化研究提供路線圖,以提高或保護產量以響應非生物逆境。本文中討論的許多進展都是在模式系統擬南芥中取得的。因此,需要深入研究,以確定作物中是否使用了類似或不同的機制。此外,很明顯,特定的細胞類型有特定的激素信號通路和輸出,因此改變細胞靶向或組織靶向的特征將需要研究這些細胞類型中的激素信號機制。此外,通過知識引導的作物新馴化來提高適應氣候變化的野生品種的產量,為納入有益的激素信號性狀提供了一條重要的新途徑。繼續深入了解植物激素在非生物脅迫不同反應中的相互作用,將對開發非生物抗逆作物具有重要意義。