晝夜節律紊亂被視為癌癥預后不良的一個標志,然而睡眠不足(SD)促進腫瘤發生的生理過程仍不明確;已有研究表明脂肪酸氧化(FAO)失調可增強腫瘤惡性特征,特別是作為FAO的限速酶,ACSL 1在癌癥中高表達,但其節律性破壞的作用未知;已知β-內啡肽,一種內源性具有類似嗎啡作用肽類物質,其晝夜振蕩與生物鐘穩態相關,具有抑制癌癥的潛能,但不同的給藥節律是否能減輕腫瘤尚待研究。
2024年5月,大連醫科大學腫瘤干細胞研究院劉強教授團隊在Cell Metabolism上發表了題為“Oncogenic fatty acid oxidation senses circadian disruption in sleep-deficiency-enhanced tumorigenesis”的研究論文近期,揭示了FAO是一種將睡眠不足與肺腫瘤發生聯系起來的晝夜節律傳感機制,并為患有睡眠不足相關癌癥的患者提供了一種β-內啡肽時間節律治療策略。

技術路線圖

FAO:脂肪酸氧化;SD:睡眠不足/睡眠剝奪;ACSL1:酰基輔酶A合成酶長鏈家族成員1;PA:棕櫚酸;PA-CoA:棕櫚酰肉堿輔酶A;CLOCK:晝夜節律穩態核心基因
研究結果
1. 脂肪酸氧化可感知睡眠剝奪導致的節律紊亂
為了探究睡眠剝奪(SD)對小鼠肺部生理節律的影響,對SD處理后不同時間點的小鼠肺組織進行組學檢測。轉錄組測序篩選出3284個在SD后喪失晝夜振蕩的基因,GO富集分析顯示這些基因在脂質代謝相關的生物過程中富集,尤其是脂肪酸氧化(FAO)。隨后的UHPLC-MS代謝組學進一步證實,SD處理使得小鼠肺組織中晝夜振蕩的脂質代謝物數量顯著減少, 47種脂質物質發生節律紊亂,其中游離脂肪酸占比最大。
以上結果表明,在基因表達和代謝功能層面,FAO均能夠感知生理條件下SD誘導的節律紊亂。
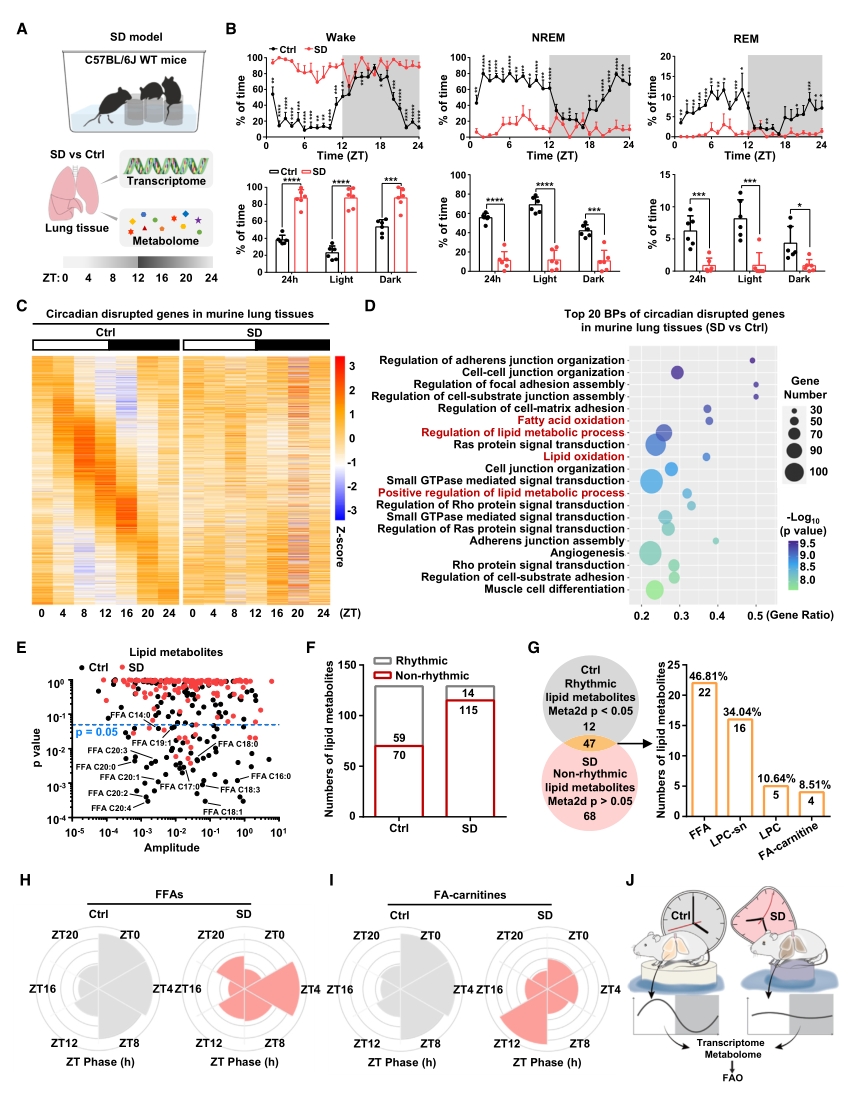
圖1. FAO可感知SD處理導致的晝夜節律紊亂
2.睡眠剝奪通過脂肪酸氧化增強癌細胞干性
在此基礎上,進一步探討SD誘導的FAO節律紊亂是否在肺癌發展中發揮作用,為此構建了肺癌小鼠模型并實施SD處理,通過CT圖像、HE染色和IHC等結果確認了肺癌形成和干細胞標志物的表達。
為明確SD促進腫瘤發生的機制,對小鼠肺癌組織進行轉錄組測序,結果顯示SD顯著上調了與FAO相關的基因表達。此外,脂質代謝組檢測也表明,SD處理后脂肪酰肉堿增加,游離脂肪酸減少,且脂肪酸氧化的最終產物乙酰輔酶A和腺苷三磷酸在肺癌中增加,并在SD條件下表現出節律紊亂。此外,向SD小鼠給予脂肪酸氧化抑制劑:依托莫司他(ETO),可逆轉肺癌進展及干細胞特性的增強。
以上結果表明,FAO感知晝夜節律紊亂是睡眠剝奪增強肺癌干細胞特性的基礎。
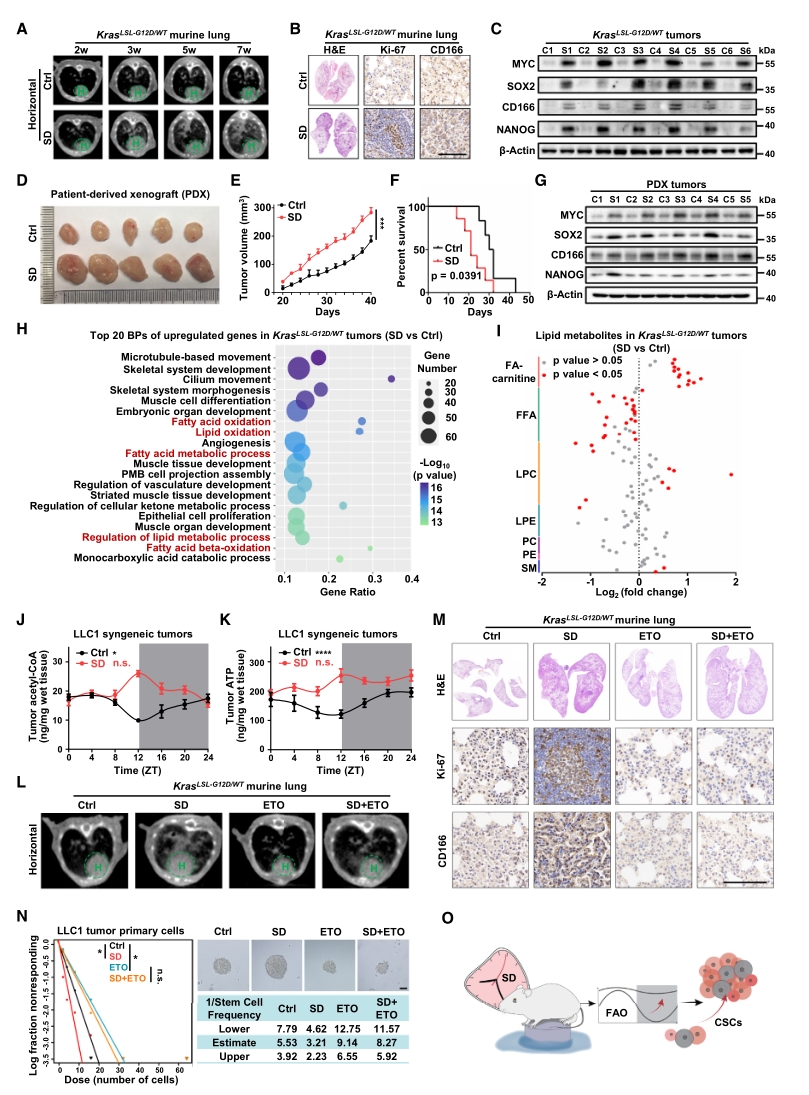
圖2. FAO節律紊亂增強SD誘導的肺癌細胞干性
3. ACSL1及其產物PA-CoA發揮重要作用
接下來尋找SD誘導FAO節律紊亂從而促進肺癌干細胞特性的關鍵分子。首先從轉錄組結果里篩選出13個同時響應節律紊亂和腫瘤進展的FAO基因,其中ACSL1在mRNA和蛋白水平均顯著上調。在小鼠體內特異性敲除該基因可逆轉SD促進的腫瘤進展,下調腫瘤組織中干細胞標志物的表達。
隨后從脂質組檢測結果里篩選可能作為ACSL1底物和產物的長鏈FAO代謝產物,發現SD可降低腫瘤組織中的棕櫚酸(PA)水平。敲低ACSL1可恢復SD誘導的PA下降,而補充棕櫚酰肉堿輔酶A(PA-CoA)——PA進入FAO的活化態,可挽救ACSL1缺失下被抑制的癌癥干細胞特性和腫瘤生長。
以上結果表明,SD擾亂ACSL1的節律,過度激活了它將PA轉化為PA-CoA的能力,從而促進FAO,最終導致腫瘤生長。
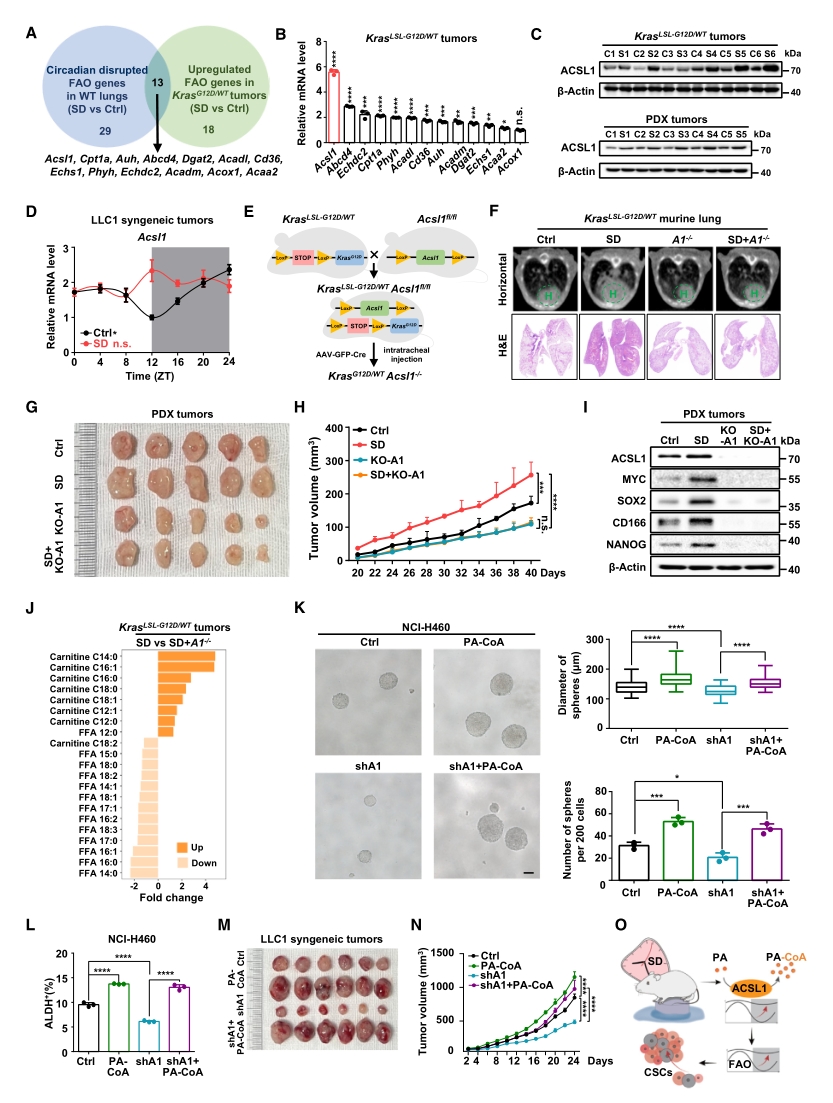
圖3. ACSL1合成的PA-CoA有助于SD誘導的肺癌細胞干性
4. ACSL1的上游調控者是CLOCK
在分子機制方面,首先在不同的細胞系中發現有兩種核心生物鐘基因:CLOCK和BMAL1,與ACSL1的表達量正相關。隨后在腫瘤模型中進行驗證,發現SD僅破壞CLOCK基因的節律振蕩,而對BMAL1無顯著影響。此外,通過分析表觀組學公共數據庫,發現CLOCK和BMAL1在多種細胞中均占據了ACSL1的啟動子區域,并且通過CHIP-qPCR和熒光素酶報告基因分析確認了結合位點。作為驗證,分別在鼠源和人源兩種腫瘤模型中敲除CLOCK基因,均觀察到了腫瘤生長和干細胞特性被抑制的現象。
以上結果表明,核心生物鐘基因CLOCK可能通過直接結合到ACSL1啟動子上的特定區域來調控該基因的表達。
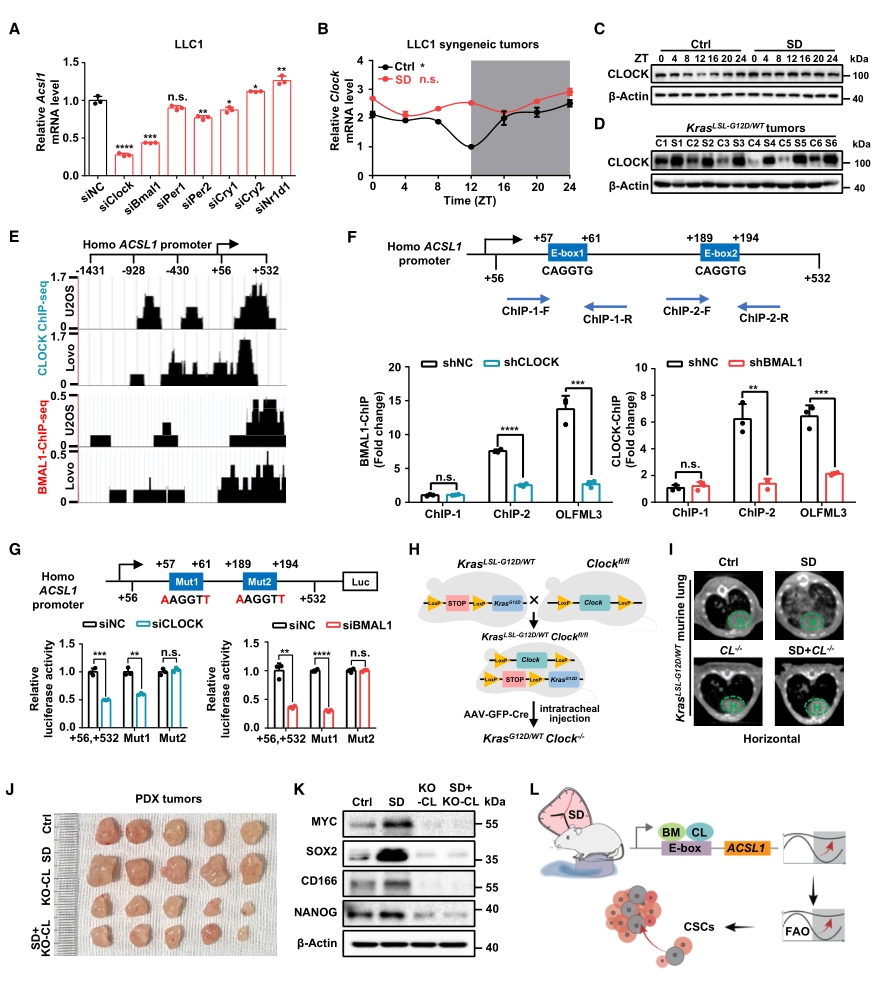
圖4. ACSL1可感知SD誘導的FAO節律失常,促進肺癌發生
5. ACSL1誘導CLOCK發生棕櫚酰化修飾,維持其穩定性
進一步的研究發現,敲低ACSL1會反過來降低CLOCK的蛋白水平,但不影響其mRNA水平,說明ACSL1可通過某些機制維持CLOCK的蛋白穩定性;同時考慮到ACSL1催化的PA-CoA可以做為蛋白質棕櫚酰化的底物,因此研究者探討了棕櫚酰化修飾在ACSL1和CLOCK互作中的作用。
首先利用酰基-生物素交換(ABE)和免疫印記等技術手段,證實了CLOCK蛋白上確實存在棕櫚酰化修飾。然后使用siRNA篩選出了23種可能的棕櫚酰轉移酶,通過敲低和過表達實驗確定了其中的ZDHHC5可介導CLOCK的棕櫚酰化。此外,還發現SD 處理一方面以ACSL1依賴的方式增強了CLOCK的棕櫚酰化修飾,另一方面也破壞了這一修飾的晝夜節律性。隨后,通過在線預測網站swisspalm鑒定了CLOCK的棕櫚酰化位點:Cys194,該位點在多物種中保守。進一步地,CLOCK蛋白的降解和泛素化實驗表明,棕櫚酰化通過抑制泛素化依賴的蛋白酶體降解來穩定CLOCK蛋白。
以上結果表明,ACSL1通過ZDHHC5使得CLOCK在Cys194上發生棕櫚酰化修飾,而這一修飾可阻止CLOCK因泛素化而降解,從而維持該蛋白的穩定性。

圖5. ACSL 1通過ZDHHC 5對CLOCK進行棕櫚酰化修飾,以此穩定CLOCK蛋白
6. 節律性的β-內啡肽給藥可作為一種治療方案
基于上述SD對腫瘤發生發展的影響,研究者考慮基于晝夜節律的內分泌治療方案。對體外培養的肺癌細胞系分別添加14種睡眠相關激素,發現β-內啡肽(Endor)可顯著下調CLOCK和ACSL1的mRNA表達水平,因此后續對β-內啡肽展開節律治療應用的研究。
對于SD條件下的荷瘤小鼠,研究人員設計了3種β-內啡肽給藥方案:每日16:00給藥(ZT8-Endor)、每日4:00給藥(ZT20-Endor)以及持續給藥(pump-Endor)。從表型來看,ZT8-Endor方案顯著逆轉了SD導致的腫瘤生長,而另外兩種方案效果不顯著。接下來使用ELISA評估了血清β-內啡肽水平,發現SD處理破壞了β-內啡肽的晝夜節律并使其保持在較低水平, 另外三種給藥處理雖都能提高β-內啡肽含量,但僅ZT8-Endor能使其晝夜節律也恢復正常。同樣地,在ACSL1和CLOCK含量和腫瘤干性標志物等指標上也存在類似的規律。
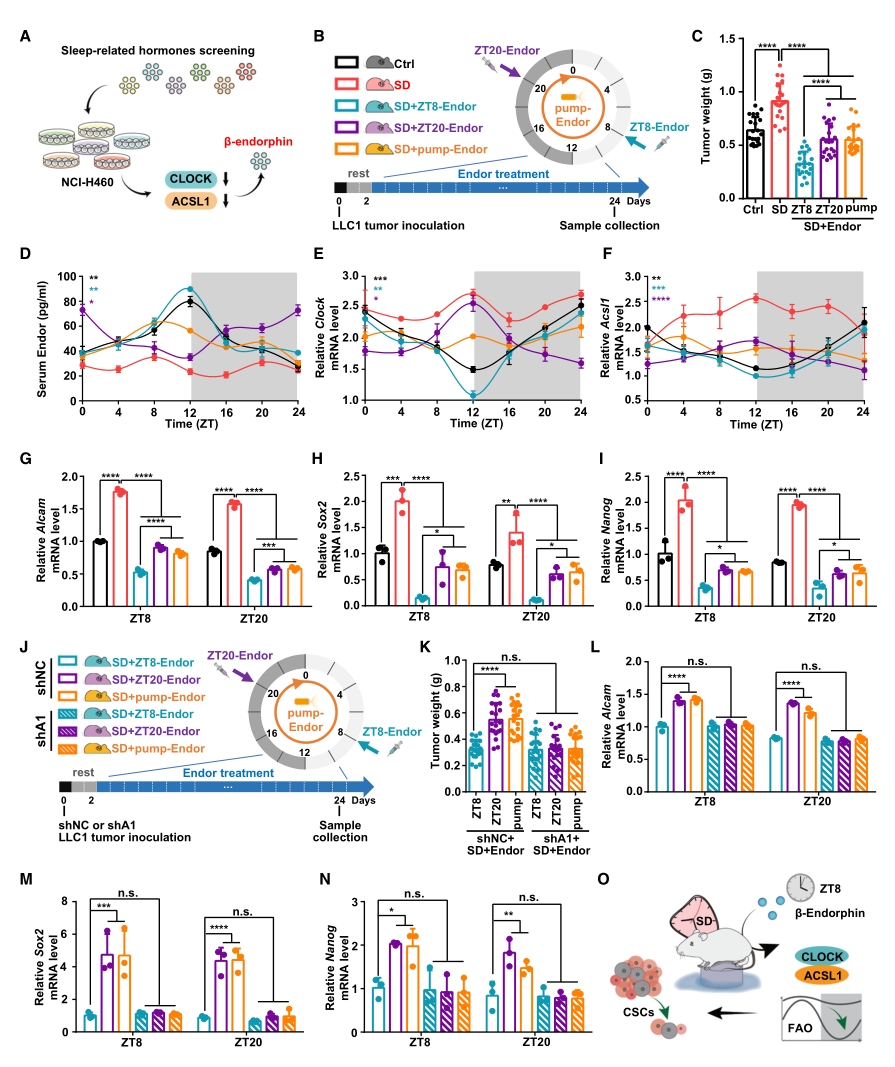
圖6. β-內啡肽可恢復受到SD干擾的CLOCK/ACSL1節律
7. 人群隊列驗證
最后為評估以上研究的臨床轉化潛力,進行了人群隊列的前瞻性和回顧性研究。肺癌患者的前瞻性隊列數據表明,血清β-內啡肽濃度與睡眠質量正相關,且是癌癥發展的潛在預測因素;既往肺癌病例的回顧性分析結果表明,血清β-內啡肽與較好的預后和生存率有關,而與CD166和Ki-67(兩種腫瘤標志物)的表達水平負相關。
總而言之,這兩組隊列數據提供了SD、β-內啡肽、CLOCK和ACSL1可能是肺癌獨立預后因素的臨床證據。
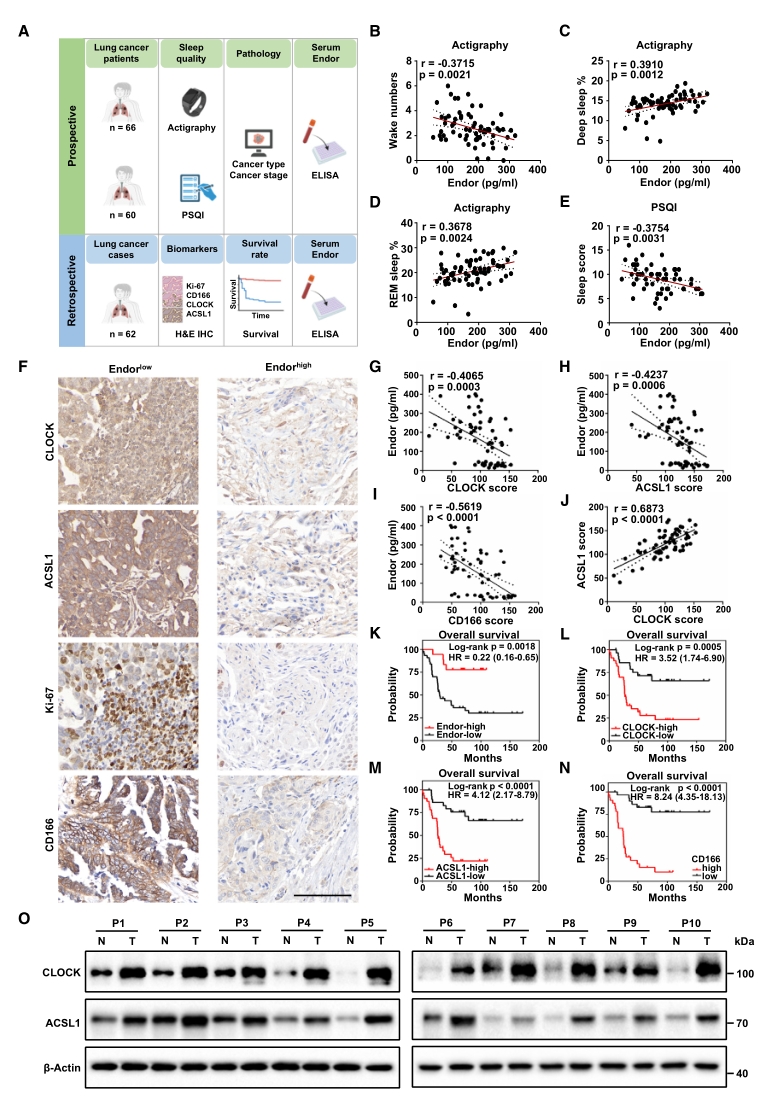
圖7. 肺癌患者的睡眠質量和β-內啡肽水平與CLOCK/ACSL1表達及預后不良呈負相關
研究結論
本研究揭示了睡眠不足(SD)與肺癌發生發展之間的潛在機制,發現了SD可導致脂肪酸氧化(FAO)紊亂,進而促進肺癌干細胞的形成和腫瘤生長。在這一過程中,ACSL1基因與CLOCK基因之間的正反饋循環起到了關鍵作用。在治療階段,通過定時給予β-內啡肽,可以有效逆轉SD引起的晝夜節律紊亂和FAO紊亂,從而減輕腫瘤的生長。此外,研究證實睡眠質量和血清β-內啡肽水平與肺癌患者的預后不良密切相關。這一發現表明,FAO作為一種晝夜節律感知機制,在連接睡眠不足與肺腫瘤進展方面具有重要意義。
參考文獻
Peng F, Lu J, Su K, et al. Oncogenic fatty acid oxidation senses circadian disruption in sleep-deficiency-enhanced tumorigenesis. Cell Metab. 2024.
請掃描二維碼閱讀原文

繪譜幫你測
麥特繪譜開創性地搭建了醫學領域高端代謝組學技術平臺,覆蓋了非靶向-全定量-代謝流等全方位的高端醫學代謝組解決方案,同時全面布局微生物組學、轉錄組學和蛋白質組學等多組學技術服務,已成為全球多組學研究者的優選合作伙伴。麥特繪譜擁有Q1000,Q500、Q300、Q200和膽汁酸、短鏈脂肪酸、色氨酸及吲哚衍生物、多胺和TMAO類等各類小分子代謝物、非靶向代謝組學和同位素示蹤代謝流技術等共40+系列檢測方法;已為數百家三甲醫院、科研院所和企業提供多組學解決方案,協助客戶與合作伙伴發表SCI文章400+篇,累計影響因子4000+,平均IF>10,包括Cell, Nature, Science, Cell Metabolism, Immunity, Gut, Hepatology, Microbiome等頂級期刊。