氨基酸是維持生命的基本單元,可相互結合生成肽進而合成蛋白質,從而介導細胞功能的發揮,包括免疫細胞。此外,氨基酸還被用于許多其他促進細胞和器官功能的過程,包括ATP的產生、核苷酸的合成以及氧化還原的平衡。免疫細胞對氨基酸有特定的需求,而生長因子刺激和T細胞激活可誘導其快速增殖,從而增加氨基酸轉運蛋白的表達,表明該過程需要氨基酸攝取的增強。來自德國馬克斯·普朗克免疫生物學和表觀遺傳學研究所的Beth Kelly和Erika L. Pearcel兩位學者總結了氨基酸代謝促進免疫細胞功能的機制,并對靶向氨基酸代謝如何調節癌癥、感染和自身免疫中的免疫反應進行了探討。相關成果發表于《Cell Metabolism》。
氨基酸的獲取與感知
每個氨基酸有其獨特的側鏈,從而具有不同的性質和功能。哺乳動物利用20種氨基酸進行蛋白質合成,當然在自然界中仍存在更多的氨基酸并影響細胞功能。這其中,9種必需氨基酸不能由人體自身充分合成而需從食物中獲取,非必需氨基酸則相反,但在人體對氨基酸的需求超過合成能力時,也會成為必需氨基酸。這種依賴發生在快速增殖的細胞中,需要氨基酸合成蛋白質和增加生物量。鑒于細胞生長和增殖的顯著變化是免疫細胞對細胞外環境變化的反應特征,對這些細胞來說,非必需氨基酸可能在免疫反應中成為條件必需的。
氨基酸獲取是細胞功能的控制點。氨基酸從外部環境攝取依賴于轉運蛋白,而在細胞內被循環利用并為其他氨基酸提供功能基團,如谷氨酰胺通過轉氨和轉酰胺作用為其他氨基酸提供氨基,蛋氨酸在蛋氨酸循環中代謝為半胱氨酸。溶酶體也含有氨基酸轉運機制,可作為細胞內氨基酸庫,對細胞功能很重要。溶酶體組氨酸轉運蛋白Slc15a4是漿細胞樣樹突狀細胞(pDCs)產生Toll樣受體(TLR)誘導的I型干擾素(IFN-I)和B細胞產生IFN-I和免疫球蛋白(Ig)G所必需的。這種溶酶體氨基酸儲存可能誘導自噬,從而促進長壽,部分是通過拮抗衰老相關CD8+T細胞功能的下降及對感染和癌癥的免疫反應。溶酶體儲存為細胞提供氨基酸庫,當蛋白質合成恢復時,這些氨基酸可以被調動起來,而自噬可能在這種情況下增加氨基酸的可用性。氨基酸可以選擇性地釋放以驅動特定的基因表達程序,從而適應長期的饑餓或營養恢復,例如通過影響表觀遺傳修飾。
氨基酸轉運蛋白調控免疫細胞功能。通過轉運蛋白攝取氨基酸是一個嚴格調控的過程,對免疫細胞的激活和功能至關重要。不同類型免疫細胞表達的轉運蛋白和受體的種類繁多,而CD4+和CD8+ T細胞的蛋白質組學分析闡明了抗原和細胞因子的刺激如何改變氨基酸轉運蛋白的表達,從而改變營養攝取,特別是激活T細胞的效應功能。與未成熟的CD8+ T細胞相比,在細胞因子如白介素(IL)-2存在的情況下通過T細胞受體(TCR)激活的CD8+ T細胞會增加細胞表面Slc1a5和Slc7a5的密度。激活的T細胞需要氨基酸的供應來支持增殖,這種激活依賴的轉運蛋白表達的增強保證了細胞可以根據需要獲取這些營養物質。轉運蛋白的表達水平也會影響T細胞功能。激活的CD4+和CD8+ T細胞表達類似的營養轉運蛋白,CD4+ T細胞的轉運蛋白拷貝數更低,營養轉運更少。這一差異,以及相比CD8+ T細胞,活化的CD4+ T細胞的核糖體水平和翻譯機制更低,可能是因為CD4+ T細胞比CD8+ T細胞的細胞質量和增殖能力更低。此外,經典和替代巨噬細胞的激活依賴于通過陽離子氨基酸轉運蛋白(CAT)2 (Slc7a2)攝取的精氨酸,而靜息狀態的巨噬細胞可能是通過CAT1 (Slc7a1)利用y+ L (Slc7a6, Slc7a7)系統攝取精氨酸。可以說先天免疫細胞的激活依賴于氨基酸轉運蛋白活性的重塑。
多種信號通路反過來控制轉運蛋白的表達。Myc是一種驅動T細胞活化的中央轉錄因子,能增加CD4+和CD8+ T細胞的Slc1a5、Slc7a1、Slc7a5、Slc38a1和Slc38a2。Myc 缺乏通過降低這些轉運蛋白的表達來阻斷活化的T細胞生長。細胞因子對T細胞生長和增殖的不同影響是由轉運蛋白的差異調控和信號通路的使用造成的。IL-2促進CD8+效應T (Teff)細胞的形成和生長,而IL-15則是體積較小的CD8+記憶T (Tmem )細胞的形成所必需的。與IL-15相比, IL-2刺激的T細胞有更高的CD98 (Slc7a5/Slc3a2)表達,并增加氨基酸攝取和蛋白質合成。這些作用依賴于IL-2培養的T細胞中持續的磷酸肌醇-3-激酶(PI3K)信號,而IL-15培養的細胞中則依賴更短暫的PI3K信號。當然仍需要更多的研究來了解細胞因子是如何通過調節氨基酸轉運蛋白從而控制免疫細胞生長的信號通路活性和動力學。
氨基酸轉運蛋白的不對稱分布影響T細胞的命運。感染期間,T細胞群體必須產生Teff細胞來對抗感染,產生Tmem 細胞提供持久的免疫,不對稱細胞分裂是T細胞實現這一目標的一種模型。在不對稱細胞分裂過程中,子細胞不平等地遺傳蛋白質、RNA和細胞器,為它們不同的命運奠定了基礎。抗原呈遞細胞(APCs)刺激未成熟T細胞的活化和增殖。基于因子(包括CD3、CD4、CD8、CD62L、轉錄因子和mTOR)的不對稱遺傳,子細胞之間,靠近APC的T細胞采用效應樣命運,而遠端T細胞采用記憶樣命運。比如Slc1a5,優先聚集在近端細胞,也增加氨基酸豐度、谷氨酰胺攝取、糖酵解、c-Myc和mTOR復合物(mTORC)1活性。這里Slc1a5對c-Myc的直接作用尚未清晰,但氨基酸的剝奪或谷氨酰胺分解的阻斷減少了子細胞之間c-Myc的差異,c-Myc水平的降低將子細胞推向記憶樣表型。同樣,Slc7a5表達升高的CD8+ T子細胞增加了CD8表達、mTORC1活性、Myc水平和糖酵解,而CD8和Slc7a5表達較低的細胞線粒體質量增加。因此,氨基酸代謝和細胞代謝特征是細胞命運的決定因素。
感知氨基酸調控免疫細胞代謝。mTORC1是一個驅動蛋白質合成、細胞生長和增殖的細胞信號中心,而GCN2激酶通過檢測空載的tRNA來感知氨基酸饑餓。mTORC1僅在資源充分時才許可免疫細胞激活。活性mTORC1促進CD8+細胞毒性T細胞(CTLs)的分化,控制Tmem細胞形成,刺激Th1和Th17的分化,而限制Treg的形成。巨噬細胞中,藥理上激活GCN2和氨基酸饑餓反應抑制IL-1β的產生以及炎性體介導的腸道炎癥。一旦被激活,先天免疫細胞和適應性免疫細胞都必須感知其氨基酸供應,以參與伴隨該激活的新的生物合成程序,在此情況下mTORC1和GCN2是氨基酸利用的主要調節者。
氨基酸支持免疫細胞代謝重編程
受體連接和細胞因子信號傳導激活免疫細胞誘導轉錄和翻譯發生劇烈變化產生細胞因子和效應分子,這需要獲得生物量來進行細胞分裂。免疫細胞重編程其代謝來支持這些異常高的新陳代謝需求。ATP形式的能量是激活的免疫細胞的主要需求,糖酵解、TCA循環和OXPHOS協同產生ATP,而氨基酸調節這些相互連接的過程。氨基酸代謝促進糖酵解和TCA循環,以及線粒體脂肪酸氧化(FAO),進一步推動線粒體代謝和ATP的產生(圖1)。
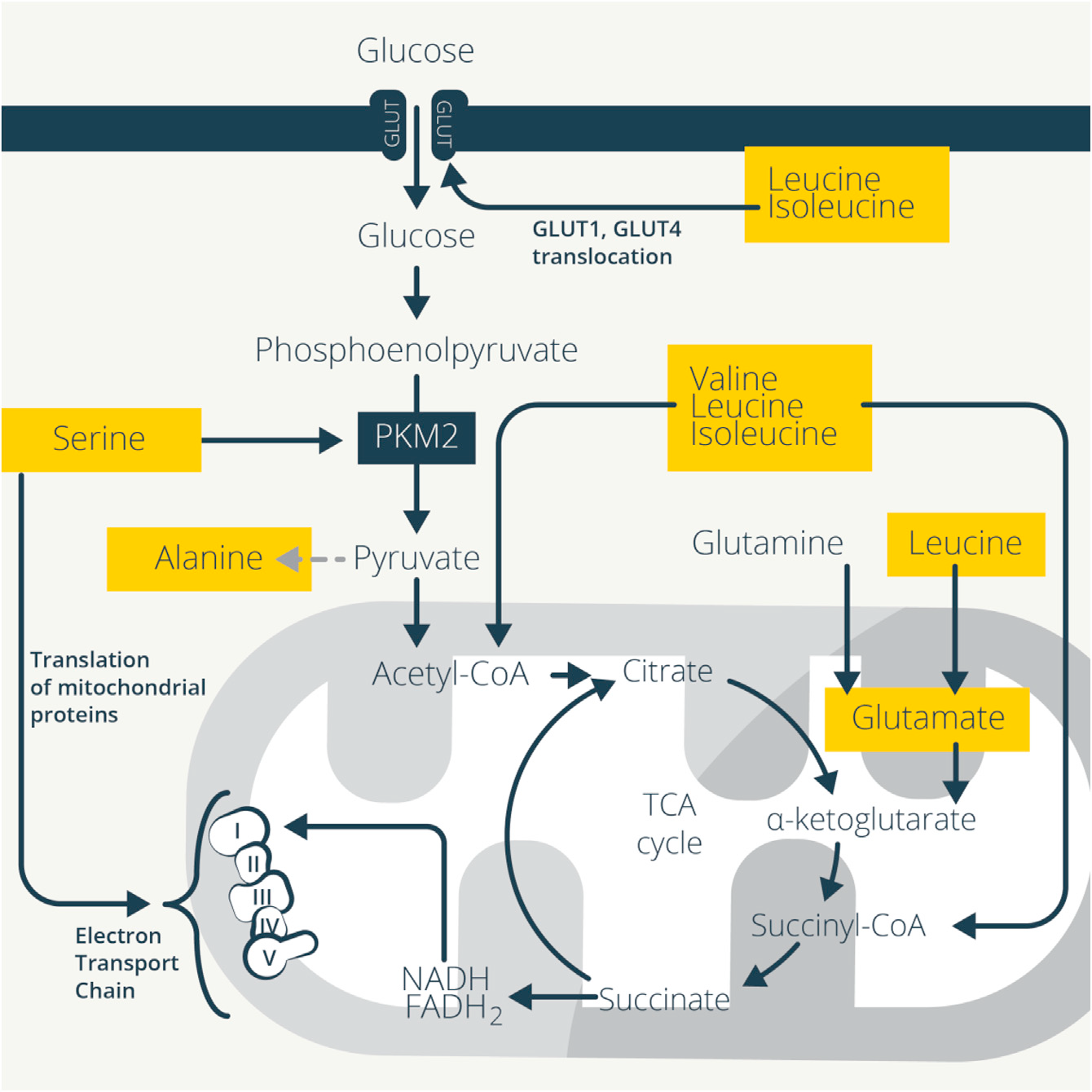
Figure 1. Amino Acids Support Glycolysis and Mitochondrial Metabolism
谷氨酰胺支持代謝重構。谷氨酰胺分解是增殖的細胞,包括激活的T細胞的主要能量產生過程。天門冬酰胺、脯氨酸或谷氨酸的補充可代謝為谷氨酰胺,但不會恢復由谷氨酰胺缺乏引起的增殖或細胞因子缺陷,表明胞外獲得而非胞內產生的谷氨酰胺是關鍵的調節因素。這解釋了為什么未成熟T細胞激活會誘導依賴于SNAT1, SNAT2 (Slc38a2) 和丙氨酸、絲氨酸、半胱氨酸偏好的轉運蛋白2 (ASCT2;Slc1a5)的谷氨酰胺攝取。盡管增殖和IL-2的產生正常,ASCT2的破壞也會抑制Th1和Th17的分化。支持T細胞分化所必需的糖酵解和線粒體代謝的增加也在Asct2-/- / CD4+ T細胞中受損,因為它們降低了葡萄糖攝取、乳酸產生和耗氧量。補充谷氨酰胺可恢復這些作用,盡管ASCT2運輸的其他氨基酸理論上也可能會導致這種表型。
谷氨酰胺支持Th1和Th17細胞的分化,但谷氨酰胺分解可能對Th17細胞更為關鍵,而谷氨酰胺的其他下游效應可能會影響Th1細胞的分化,這在缺乏谷氨酰胺酶1 (Gls1)的小鼠身上得到了證明。Gls1缺乏限制αKG的供應,損害Th17細胞的分化。Gls1中斷抑制mTORC1和IL-2信號,進而抑制T細胞分化。同時,Gls1缺乏增加轉錄因子T-bet,促進Th1和CD8+ CTLs的分化和效應功能。限制谷氨酰胺分解可能增加其他過程的谷氨酰胺有效性,或者可能限制αKG產生支持T-bet介導的Th1細胞分化和功能的表觀遺傳特征。治療上,Gls1抑制可改善實驗性自身免疫腦脊髓炎(EAE)和類風濕關節炎模型的高炎癥。谷氨酰胺分解是支持特定T細胞亞群功能的主要分解代謝途徑。
不同免疫細胞類型具有不同的谷氨酰胺代謝。IL-2/IL -12誘導的Myc依賴性自然殺傷(NK)細胞的激活需要Slc7a5介導的谷氨酰胺攝取。這種情況下谷氨酰胺的獲取而非降解是重要的,因為是谷氨酰胺剝奪而不是谷氨酰胺分解的阻斷抑制NK細胞的激活。相反,谷氨酰胺分解支持激活的B細胞產生抗體。激活的B細胞中let-7adf microRNA簇過表達抑制ASCT2和Gls1的表達,減少IgM和IgG的產生。
谷氨酰胺對αKG的回補在促炎的M1樣巨噬細胞中增加,并且TCA循環的中斷導致琥珀酸的積累,驅動IL-1β的產生。一氧化氮(NO)介導的丙酮酸脫氫酶和烏頭酸酶-2抑制限制了代謝產物進入TCA循環,促進炎癥巨噬細胞中谷氨酰胺回補為αKG。谷氨酰胺流入TCA循環是IL-4誘導M2樣巨噬細胞極化的原因。漿細胞也使用谷氨酰胺來替代缺乏的谷氨酸和天門冬氨酸。相比之下,活化的NK細胞使用谷氨酰胺來補充TCA循環中間產物并增加OXPHOS,但葡萄糖供應的檸檬酸-蘋果酸穿梭在這些細胞中對TCA循環活性的驅動作用更大。NKT細胞兼具先天和適應性免疫細胞的特征,并且表達高度受限的TCR庫,也需要谷氨酰胺來生存和增殖。很明顯,許多免疫細胞增加了谷氨酰胺的攝取。谷氨酰胺分解支持其中一些免疫細胞的功能,而另一些則將谷氨酰胺導入其他代謝途徑。增強對不同細胞類型中谷氨酰胺利用的了解,將使谷氨酰胺代謝在治療中有更明確的靶向作用。
支鏈氨基酸(BCAAs)為TCA循環提供中間產物,乙酰CoA和琥珀酰CoA。亮氨酸轉氨也會產生谷氨酸,通過αKG補充TCA循環。與谷氨酰胺轉運蛋白類似,免疫細胞激活后,許多中性氨基酸轉運蛋白增加。L型氨基酸轉運蛋白1(LAT1;Slc7a5)很好地說明了氨基酸轉運蛋白如何將病原體感染與免疫反應聯系起來。在體內,李斯特菌感染增加了Slc7a5在T細胞的表達,而IL-2保持了這種表達,維持了對活化T細胞的持續BCAA供應。抑制Slc7a5阻礙IFN-γ和IL-17的生成以及Th1和 Th17細胞的發育,但Treg細胞發育正常。缺乏Slc7a5的T細胞不能經歷激活所必需的mTORC1和Myc依賴的糖酵解增加。特別是,單獨敲除Slc7a5很大程度上復制了Myc缺乏對活化的CD4+T細胞蛋白質組的影響,并損害T細胞的生長。單一轉運蛋白的缺失具有強烈的影響,說明氨基酸轉運和獲取對T細胞功能的重要性。
Slc7a5也會引起巨噬細胞代謝重整。LPS增加巨噬細胞中BCAA轉運蛋白的表達,并且缺乏Slc7a5介導的亮氨酸轉運的巨噬細胞糖酵解和IL-1β的產生降低。CD98在分泌抗體的漿細胞中的表達較高,其缺失會影響抗體的產生。細胞表面CD98的表達與漿細胞壽命有關,高表面CD98水平的漿細胞比低表面CD98表達的短壽命漿細胞分泌更多的抗體。有趣的是,這些有著不同壽命的群體具有相似的轉錄譜,但代謝參數如CD98表達、葡萄糖攝取以及丙酮酸依賴的線粒體呼吸更能區分壽命差異的漿細胞。因此,操縱漿細胞代謝是增加體液免疫對病原體的持續時間的潛在有效的方法。
BCAA轉氨酶(BCAT1)可將BCAA轉化為支鏈α酮酸,鏈α酮酸再脫羧形成CoA衍生物。抑制BCAT1可限制糖酵解和耗氧量,以及抗炎物質衣康酸的產生。早期研究顯示,喂養BCAAs缺失飲食小鼠產生缺陷抗體和細胞毒性T細胞反應,以響應鼠傷寒沙門氏菌或乳腺癌,而補充BCAAs可增強肝炎小鼠模型的肝臟CD8+ T活性,也可增加術后創傷或膿毒癥患者淋巴細胞數量和對皮膚抗原的反應。纈氨酸促進肝硬化患者DC IL-12的產生。BCAA還可通過刺激葡萄糖攝取促進糖酵解以支持免疫細胞的代謝重編程。大鼠肌肉細胞中,亮氨酸和異亮氨酸可能依賴于PI3K和蛋白激酶C增加葡萄糖轉運蛋白GLUT1和GLUT4向細胞表面的易位,以增加葡萄糖攝取。亮氨酸提供的乙酰CoA也可以乙酰化和激活mTORC1,進而增強糖酵解。
絲氨酸促進糖酵解,它連接和異構激活丙酮酸激酶M2(PKM2),在糖酵解最后一步將磷酸烯醇丙酮酸轉化為丙酮酸。LPS誘導巨噬細胞中PKM2驅動代謝向糖酵解轉變和IL-1β誘導。CD4+ T細胞的TCR刺激增加PKM2的核易位,這增強糖酵解以支持Th1和Th17細胞的生成、腫瘤壞死因子(TNF)和IL-17的產生。絲氨酸限制也抑制PKM2以減少動脈粥樣硬化中巨噬細胞的過度激活,并且抑制絲氨酸合成可降低LPS誘導的內毒素血癥中IL-1b和TNF的產生。
絲氨酸也促進線粒體代謝。線粒體翻譯和呼吸作用需要絲氨酸羥甲基轉移酶(Shmt2),Shmt2缺失的小鼠具有呼吸障礙,這種效應依賴于來自絲氨酸的一碳(1-C)單位的生成。線粒體翻譯起始依賴于由絲氨酸通過Shmt2形成的修飾tRNA,N-甲酰甲硫氨酰tRNA(fMet-tRNAMet)。1-C單元的補充恢復了Shmt2缺乏的Jurkat細胞呼吸和線粒體翻譯的缺陷。這條途徑似乎在低糖條件下特別重要,可誘導細胞以補償方式增加線粒體代謝,因此,它也可能對免疫細胞從糖酵解轉向線粒體代謝很重要。然而,即使是有足夠葡萄糖支持糖酵解增加的T細胞,OXPHOS和效應功能也依賴于絲氨酸下游的1-C代謝。甲基四氫葉酸脫氫酶2催化線粒體fMet-tRNAMet形成的中間步驟,生成NADH。在線粒體呼吸受損的胰腺組織中,這種NADH可以積累到有毒水平,從而抑制細胞生長。
雖然其他氨基酸可以為糖酵解和TCA循環提供原料,但有時T細胞并不用它們進行這些途徑。胞外丙氨酸對T細胞早期激活是必需的,盡管丙氨酸可代謝為糖酵解的終產物丙酮酸,但它并不用于激活T細胞,反而用于蛋白質的合成。此外,雖然丙氨酸可以通過丙氨酸轉氨酶從葡萄糖合成,但這種酶在激活早期的T細胞中的表達較低,細胞轉而通過SNAT1吸收胞外丙氨酸。這保留了丙酮酸以供應乙酰CoA、TCA循環活性和OXPHOS,而胞外丙氨酸用于蛋白質合成。
氨基酸控制硫和氧化還原代謝
免疫細胞的激活往往取決于ROS的增加,LPS誘導巨噬細胞產生ROS從而驅動細胞因子的產生和殺菌。ROS可以激活活化的T細胞(NFAT)的核因子,增加T細胞的增殖。對未成熟B細胞的BCR刺激也會引起大量的ROS產生和鈣誘導的線粒體功能障礙,這會導致激活誘導的細胞死亡,除非B細胞也接收了共刺激信號,如TLR或CD40參與。因此,細胞有多種抗氧化機制來控制ROS水平,氨基酸對于維持這些防御和氧化還原平衡至關重要(圖2)。
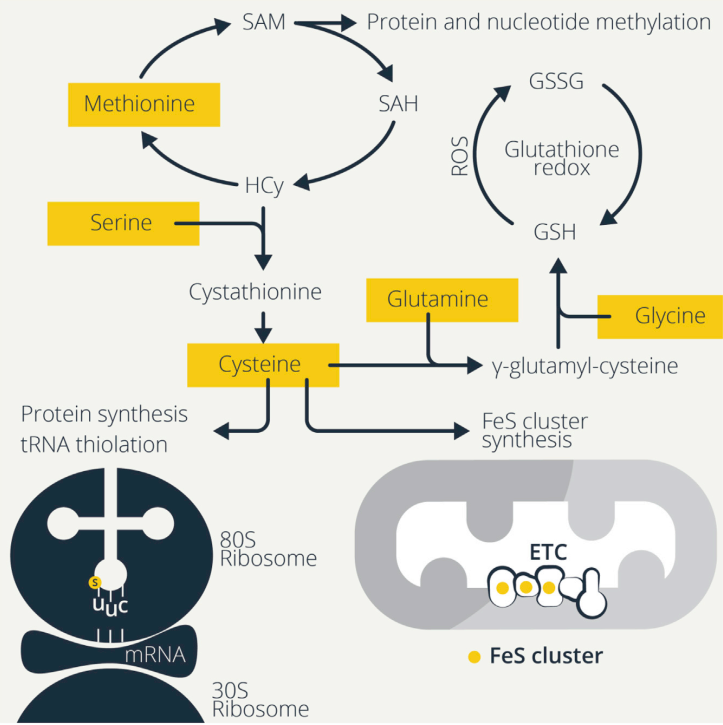
Figure2. Sulfurous Amino Acids Maintain Redox Balance and Drive Protein Synthesis
谷胱甘肽是T細胞和巨噬細胞功能所必需的。谷胱甘肽由甘氨酸、谷氨酸和半胱氨酸組成,因此,這些氨基酸的供應決定了谷胱甘肽水平。還原型谷胱甘肽(GSH)被氧化成為氧化谷胱甘肽(GSSG),谷胱甘肽還原酶(GSR)將GSSG重新還原為GSH,保持細胞內ROS解毒的GSH池。免疫細胞激活后ROS升高,增加了對GSH的需求,這就需要其組成氨基酸的供應。T細胞激活時GSH水平確實會增加,并且GSH從頭合成而非GSSG循環對T細胞分化和功能的發揮具有重要作用。此外,谷胱甘肽還影響T細胞活化過程中糖酵解和谷氨酰胺利用率的增加。有條件的Gclc缺失消除了mTORC1的激活、NFAT的核積累以及激活的CD4+和CD8+T細胞中Myc依賴的向糖酵解和谷氨酰胺分解的代謝轉變。這些細胞最初正常激活,但不能滿足擴展的代謝需求而無法增殖。相反,GSH合成受損時,絲氨酸激活mTOR,從而抑制Foxp3的表達。絲氨酸限制可恢復Foxp3的表達和Treg細胞抑制活性,降低體內致死性自身炎癥。因此,Treg細胞中GSH的一個功能是限制絲氨酸代謝,從而維持其抑制功能。巨噬細胞也利用絲氨酸為GSH產生甘氨酸,這是LPS誘導的IL-1βmRNA表達所必需的。這種絲氨酸-甘氨酸-GSH調節回路說明了氨基酸和小肽如何調節其他氨基酸的代謝。
半胱氨酸/胱氨酸導入部分通過促進GSH的合成支持免疫細胞功能。靜息B細胞不表達xCT(Slc7a11),但激活后,增加xCT表達和胱氨酸的導入,可能是為產生GSH來對抗ROS的產生,否則會導致激活誘導的細胞死亡。DCs的LPS激活增加了系統XC-胱氨酸-谷氨酸抗逆向轉運活性,這可能需要增加GSH的合成以對抗激活后ROS的增加。DCs還可以輸出GSH來塑造細胞外氧化還原環境,特別是在DCs和T細胞之間的免疫突觸周圍。GSH分解為半胱氨酸,降低細胞外氧化還原電位以及促進CD4+ Teff細胞增殖,可能是因為CD4+ Teff細胞獲得半胱氨酸用于GSH的產生和隨后的增殖。此外,Treg細胞與DCs和CD4+ Teff細胞共培養耗盡細胞外半胱氨酸,這可能解釋了Treg細胞對Teff細胞增殖的抑制作用。因此,天然免疫細胞調節氨基酸以調控適應性免疫細胞的功能。
半胱氨酸支持硫依賴的代謝。半胱氨酸是GSH功能發揮的關鍵氨基酸,因為硫的供應。半胱氨酸主要以其氧化形式,胱氨酸,存在于氧化細胞外環境中。胱氨酸的攝取對T細胞的激活、增殖和DNA合成是必需的。阻斷T細胞對胱氨酸的攝取對EAE有保護作用。與B細胞相似,靜息T細胞不表達xCT,但激活后表達xCT以促進胱氨酸的攝取。當胞外半胱氨酸供應有限時,蛋氨酸向半胱氨酸的轉硫維持半胱氨酸依賴的過程。
半胱氨酸的硫供應對于通過合成鐵硫(FeS)簇來維持線粒體代謝也是至關重要的。半胱氨酸脫硫酶NFS1從半胱氨酸中去除FeS簇的硫,并且半胱氨酸限制會降低FeS簇的合成。失去FeS簇本身會影響代謝途徑,因為含有FeS簇的烏頭酸酶不能再代謝檸檬酸,從而導致這種TCA循環中間體積累以及用于脂肪酸合成。精氨酸來源的NO破壞FeS的穩定性,表明NO可能拮抗M1樣巨噬細胞線粒體代謝。此外,FeS簇合成的降低還會導致游離鐵的過量,導致細胞鐵死亡。胱氨酸-GSH通訊可調節氨基酸信號傳導和鐵死亡。
NFS1也會利用半胱氨酸的硫原子來生成硫醇化tRNA。賴氨酸、谷氨酰胺和谷氨酸tRNAs可以在擺動位點(U34)的尿苷酸上硫醇化,通過核糖體幫助tRNA易位來促進翻譯。在硫限制,如蛋氨酸和半胱氨酸的限制過程中,這種tRNA硫醇化降低,在營養受限時翻譯減弱。免疫細胞中,FeS簇合成與tRNA硫醇化尚未研究,但在這里可能會發揮作用,因為線粒體代謝的維持和翻譯的增強是免疫細胞活化的基礎。
參考文獻
Kelly and Pearce, Amino Assets: How Amino Acids Support Immunity. Cell Metabolism (2020). https://doi.org/10.1016/j.cmet.2020.06.010.
原文閱讀,請長按識別下方二維碼

精彩推薦
2. Geroscience︱代謝組-微生物組-免疫因子關聯分析揭示大鼠機體隨年齡變化規律
3. Nature Reviews Clinical Oncology | 代謝調節提高免疫治療
4. Immunity | 重磅綜述: 飲食-菌群-免疫互作
5. TEM | 宿主與菌群互作的信號分子--AAA相關菌群代謝物
6. Cell子刊 | 癌細胞并不孤單--腫瘤微環境中的代謝通信
