細胞黏附分子(CAM)在建立和維持突觸連接中至關重要。最新證據表明,突觸間隙內的一些分泌因子,包括C1q樣蛋白,通過連接兩側的CAM在突觸前和突觸后起著重要的橋接作用。然而,這些分泌因子在突觸組裝中的作用機制仍未完全闡明。2025年12月,南方科技大學生命科學學院神經生物系魏志毅副教授課題組聯合深圳灣實驗室/北京大學深圳研究生院張勃課題組在Nature Communications (IF 15.7) 期刊發表題為“Structural basis of calcium-dependent C1ql1/BAI3 assemblies in synaptic connectivity”的文章。研究聚焦 C1ql1與突觸后膜受體BAI3相互作用介導突觸連接的分子機制,揭示了鈣調控下C1ql1的動態組裝及其對突觸形成與維持的關鍵作用,為理解突觸連接的分子基礎提供了重要見解。

· 維真助力 - 腺相關病毒·
實驗動物
P21 C1ql1-KO 小鼠
病毒產品
AAV2/9-CAG-eGFP-WPREpA;
AAV2/9-CAG-HA-mC1ql1-T2A-eGFP-WPRE-pA;
AAV2/9-CAG-HA-mC1ql1(A209S)-T2A-eGFP-WPREpA;
AAV2/9-CAG-HA-mC1ql1(D208N)-T2A-eGFPWPRE-pA;
AAV2/9-CAG-HA-mC1ql1(Y247A)-T2AeGFP-WPRE-pA;
注射方式及部位
腦立體定位注射雙側下橄欖核(IO)
注射量
250 nL/site,總500nL

IO中AAV病毒注射的共聚焦成像
研究結果
1、C1ql1_gC1q以鈣離子依賴的方式與BAI3的C末端擴展CUB結構域相互作用
C1ql蛋白共享相似的結構域組織,由NT區、中間膠原樣(CL)序列和C末端典型的gC1q結構域組成。研究團隊通過分析型尺寸排阻色譜和多角度光散射發現,gC1q結構域在溶液中存在三聚體和六聚體兩種組裝形式,二者可動態轉換。X射線晶體結構顯示,六聚體由兩個三聚體以“尾對尾”方式結合形成,每個三聚體中的β4-β5發夾結構外翻,與另一個三聚體的對應發夾結構交換,形成結構域交換界面。這一過程與中央鈣離子軸的部分重組直接偶聯。BAI3被認為是PC突觸后膜上的特異性CAM,與CF終末釋放的C1ql1相互作用,介導CF-PC突觸的發育和突觸傳遞。通過截斷體結合實驗確定,BAI3中與C1ql1_gC1q結合的最小區域是CUB結構域,尤其是eCUB²?³片段表現出最高親和力。進一步截除延伸序列會導致結合完全喪失,說明CUB結構域后的延伸序列對互作至關重要。ITC與aSEC分析表明,在鈣離子濃度低于0.1 mM時結合顯著減弱,而在≥0.5 mM時形成穩定復合物。這表明生理水平的突觸間隙鈣濃度是保證C1ql1與BAI3有效互作的關鍵條件。此外,gC1q三聚體與六聚體均能以鈣依賴方式結合BAI3 eCUB,但六聚體形成穩定復合物所需鈣離子濃度更高。此外,BAI3的結合會阻斷gC1q三聚體與六聚體之間的動態轉換。

C1ql1_gC1q以Ca2+依賴的方式與BAI3_eCUB相互作用
2、C1ql1_gC1q/BAI3_eCUB復合物的冷凍電鏡結構
研究團隊通過冷凍電鏡技術解析了C1ql1_gC1q六聚體與BAI3 eCUB復合物的三維結構,整體分辨率約為3.5 Å。結構顯示一個gC1q六聚體結合四個BAI3 eCUB分子,每個三聚體單元結合兩個eCUB。界面處存在一個關鍵的“橋接”鈣離子,同時與gC1q的D193、D235及eCUB的D63配位,直接穩定了相互作用網絡。結合界面的關鍵殘基(如gC1q的Y247與eCUB的T65、K66等)通過氫鍵與疏水作用進一步加固了復合物組裝,相關突變實驗證實了這些結構觀察到的相互作用對于C1ql1/BAI3結合至關重要。結合特異性方面,C1ql 家族成員均能與BAI2/3結合,但不與BAI1結合,源于BAI1中 E60/T65 殘基不保守。進一步的研究顯示gC1q三聚體/六聚體轉換有助于C1ql1與細胞表面的BAI3結合。全長C1ql1通過 N 端(NT)介導的二硫鍵連接和gC1q結構域介導的結構域交換,共同形成高階寡聚體,該組裝模式對其與BAI3的結合及突觸功能至關重要。
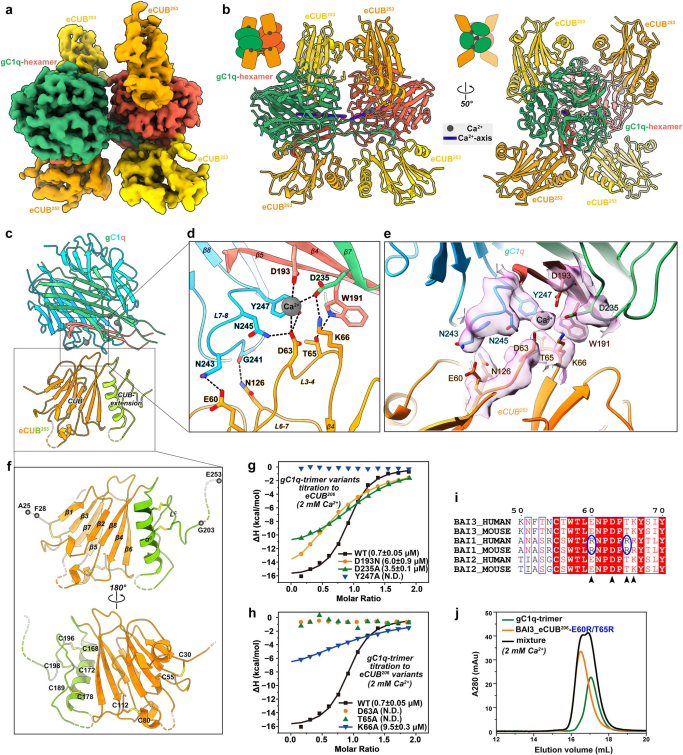
gC1q/eCUB相互作用的結構和生化分析
3、C1ql1的動態組裝及其與BAI3的相互作用對于維持攀爬纖維突觸至關重要
先前的工作報道了C1ql蛋白影響培養的海馬神經元中的突觸密度,表明gC1q的不同寡聚體形式可能對突觸性質具有不同的影響。因此研究團隊探討了gC1q六聚體對體內突觸,特別是CF突觸的作用。在C1ql1敲除小鼠中,蒲肯野細胞近端與遠端樹突上的vGluT2陽性突觸密度分別降低約30%和60%,且突觸斑塊尺寸異常增大,表明C1ql1對CF-PC突觸的維持具有關鍵作用。進一步在C1ql1敲除小鼠中利用AAV病毒載體表達野生型及突變型C1ql1進行挽救實驗,發現野生型C1ql1可部分恢復vGluT2突觸密度,破壞BAI3結合的Y247A突變體無法挽救突觸丟失;干擾gC1q三聚體/六聚體動態轉換的突變體(D208N、A209S)同樣無法恢復突觸數量,其表型與結合缺陷突變體類似。以上結果證明,gC1q結構域的三聚體/六聚體動態轉換能力,以及C1ql1與BAI3的有效結合,兩者共同構成維持CF-PC突觸完整性的必要條件。
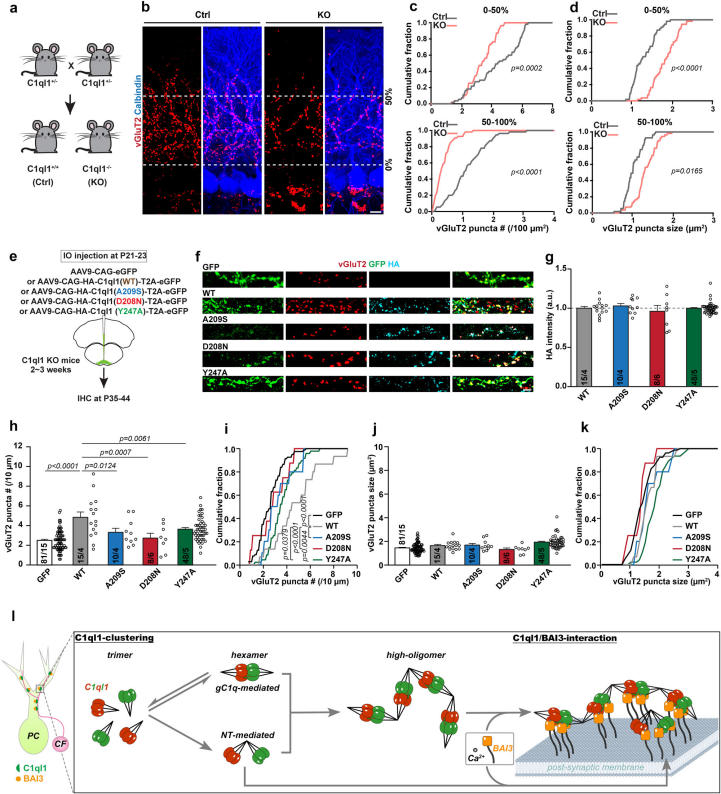
C1ql1的gC1q六聚體是維持CF-PC突觸數量所必需的
研究結論
本研究發現C1ql1的三聚體球狀 C1q(gC1q)結構域會發生鈣調控的結構域交換事件,進而形成六聚體。鈣離子對 C1ql1_gC1q 六聚體與BAI3的延伸 CUB 結構域形成的復合物具有穩定作用。借助gC1q六聚體,全長C1ql1可進一步組裝成線性簇,這可能為BAI3 受體在細胞膜上的聚集提供支架。細胞實驗和體內研究證實,gC1q介導的C1ql1動態組裝在受體聚集和突觸維持中發揮作用。綜上,研究結果揭示了一種合理的分泌因子介導突觸連接的機制,該機制由鈣調控的C1qls組裝及其與細胞粘附分子的相互作用驅動。