
期刊:Cell discovery
影響因子:13.0
主要技術(shù):scRNA-seq、scATAC-seq
導(dǎo)語
人類小腦包含多種神經(jīng)元,與大腦的發(fā)育模式完全不同。本研究通過scRNA-seq,scATAC-seq 和空間轉(zhuǎn)錄組學(xué)分析了孕周(GW)13-18周的胎兒樣本,探索發(fā)展中的人類小腦細胞多樣性和發(fā)育程序。作者鑒定了跨物種保守的暫時性顆粒細胞祖細胞,并從多個角度對顆粒細胞(GC)和浦肯野細胞的特殊模式進行了解剖,并在凌唇中發(fā)現(xiàn)了一個新的潛在神經(jīng)上皮細胞群。此外,作者多角度對比了各神經(jīng)亞型的物種保守性。最后,作者解析了浦肯野細胞和單極刷狀細胞的各亞型的分子特征,并揭示了控制其多樣化的基因調(diào)控網(wǎng)絡(luò)。本研究提供了一個人胎兒小腦的多組學(xué)圖譜,加深了我們對人類小腦的發(fā)育和空間組織的理解。
主要技術(shù)
scRNA-seq、scATAC-seq
主要結(jié)果
1. 發(fā)育中的人類小腦的多組學(xué)分析
作者選取GW13-18周的胎兒小腦樣本進行scRNA-seq、空間轉(zhuǎn)錄組學(xué)分析(Stereo-seq)以及scATAC-seq。從62,247個高質(zhì)量細胞中鑒定了GABA能神經(jīng)元(如中間神經(jīng)元、浦肯野前體細胞)、興奮性譜系細胞(如未成熟顆粒細胞、顆粒細胞祖細胞)以及HES1+祖細胞和膠質(zhì)細胞,并識別了各細胞類型的特異性標(biāo)記基因。RNA速率分析揭示了祖細胞分化軌跡,ATOH1+興奮性祖細胞(顆粒細胞系)在增殖期高表達G2/M期標(biāo)志物(如TOP2A),而PTF1A+祖細胞比例較低;浦肯野前體細胞(PKCs)在GW13占比最高,隨后迅速減少,而未成熟顆粒細胞(UBCs)占比在GW16達到峰值。空間轉(zhuǎn)錄組學(xué)顯示GW13樣本中細胞類型呈多層環(huán)狀分布(如外顆粒層、內(nèi)顆粒層),GW16樣本中可區(qū)分菱唇、浦肯野細胞層及核團區(qū)域,與解剖結(jié)構(gòu)一致。scATAC-seq與scRNA-seq數(shù)據(jù)整合分析表明染色質(zhì)可及性與基因表達譜高度相關(guān)。
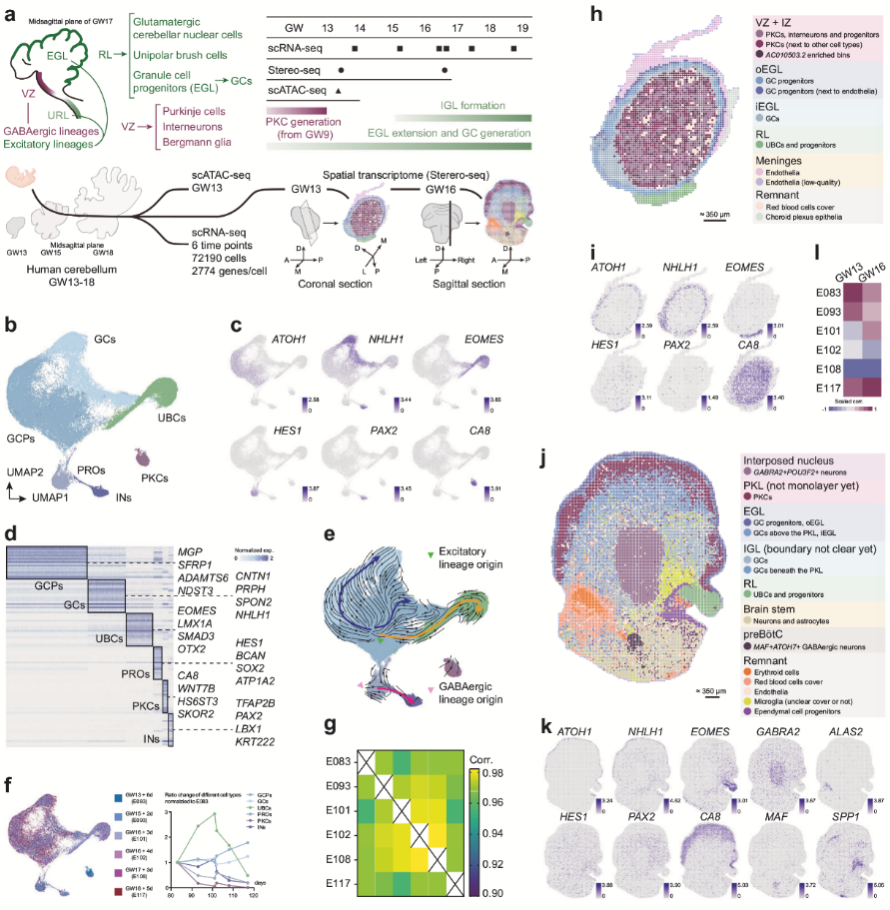
圖1 人類發(fā)育小腦的多組學(xué)分析
2. GC譜系的發(fā)育軌跡和瞬時擴增祖細胞
NRN1+小腦興奮性神經(jīng)元中,GC占比最高,隨后作者從多方面解析了GC的發(fā)育。通過細胞周期評分和擬時序分析,作者將GC譜系分為 增殖期祖細胞(AT+GCPs和ND+GCPs) 和后分裂成熟GCs。其中ND+GCPs(NEUROD1+祖細胞)具有增殖特征(表達S/G2/M期標(biāo)志物),與AT+GCPs相比不表達SOX2,高表達ELAVL2,且在小鼠中具有跨物種保守性。NEUROD1 的表達早于其他成熟GC標(biāo)記基因,可能在早期驅(qū)動分化。空間轉(zhuǎn)錄組數(shù)據(jù)揭示了 ND+GCPs與AT+GCPs的“內(nèi)-外”分層分布(iEGL和oEGL),與小鼠中瞬時擴增祖細胞的空間定位一致。后分裂GCs特異性表達 PRPH(神經(jīng)元中間絲蛋白),僅在iEGL表達,可作為iEGL標(biāo)志物;而 SFRP1 與PRPH表達互補,提示功能分化。CPLX3(前iEGL特異性)和 SV2B(iEGL富集)參與神經(jīng)遞質(zhì)分泌調(diào)控,可能與PRPH形成蛋白互作網(wǎng)絡(luò),協(xié)同調(diào)控GC遷移。整合多組學(xué)構(gòu)建基因調(diào)控網(wǎng)絡(luò),鑒定出 ATOH1、YBX3和MEF2C 等關(guān)鍵轉(zhuǎn)錄因子。 ND+GCPs中 Ras/Rap1信號通路 和趨化因子激活,調(diào)控神經(jīng)分化形態(tài)變化,且該通路在人類和小鼠中保守。
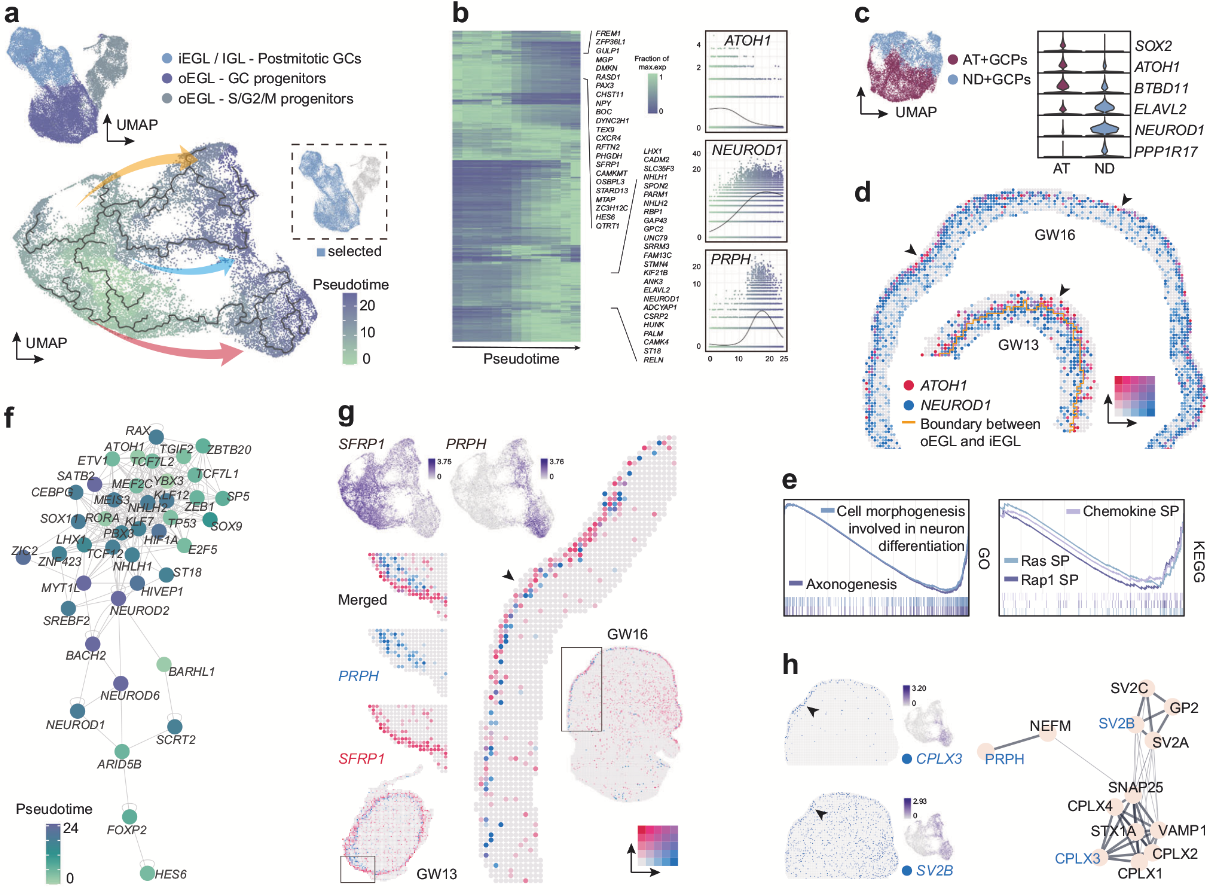
圖2. GC亞群發(fā)育軌跡和EGL祖細胞亞型分析
3. GC亞群特征及其空間分布
小腦不同區(qū)域(如前葉、后葉、絨球小結(jié)葉)表現(xiàn)出獨特的基因表達模式。例如,BARHL1和TLX3在顆粒細胞祖細胞中互斥表達,分別在小腦前部和后部區(qū)域高表達。空轉(zhuǎn)數(shù)據(jù)顯示,EBF2和HEY1富集于背內(nèi)側(cè)區(qū)域(可能與蚓部相關(guān)),而PRR35在腹外側(cè)區(qū)域(半球)表達,揭示了顆粒細胞在前后軸(A-P)和背內(nèi)側(cè)-腹外側(cè)軸(DM-VL)的異質(zhì)性。單細胞數(shù)據(jù)顯示,TLX3在人類后葉和絨球小結(jié)葉均表達,但在小鼠中僅在后葉出現(xiàn);PARM1在人類后葉和小鼠前葉富集,呈現(xiàn)相反的分布模式。這種差異提示小腦發(fā)育的分子機制可能存在物種特異性演化。整合單細胞多組學(xué)數(shù)據(jù)(scRNA-seq和scATAC-seq)發(fā)現(xiàn),A-P軸分布受特定轉(zhuǎn)錄因子調(diào)控,這些因子可能通過調(diào)控增強子活性驅(qū)動區(qū)域特異性基因表達。
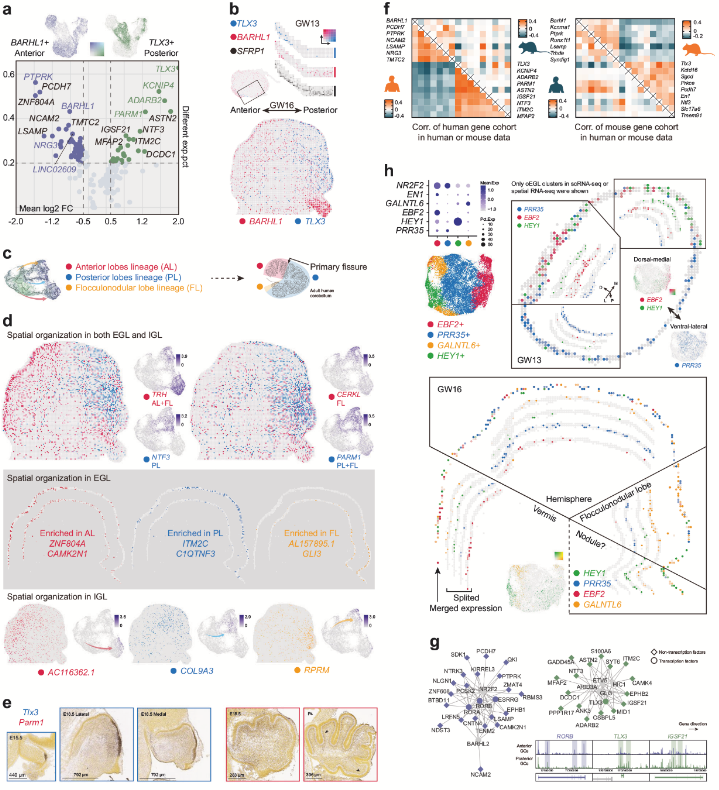
圖3 GC 亞群的空間分布和基因表達特征
4. UBC譜系發(fā)育
UBC和GC都被認為是從菱形唇(RL)中的WLS+祖細胞產(chǎn)生的神經(jīng)元譜系。作者鑒定出一類新型潛在神經(jīng)上皮細胞群(UBCpro-1),高表達COL2A1、PIK3C2G及干性標(biāo)記物(如SOX2、CYP26B1)。空轉(zhuǎn)數(shù)據(jù)顯示,COL2A1高表達于最內(nèi)側(cè)區(qū)域,與CALCB(RL腹側(cè)區(qū)標(biāo)記)和EOMES(RL亞腦室區(qū)標(biāo)記)形成“由內(nèi)向外”的分布模式,作者推測UBCpro-1可能分化為經(jīng)典的WLS+CALCB+ UBC祖細胞(UBCpro-2)。此外,作者還發(fā)現(xiàn)了UBC的兩種潛在亞型:1. LMX1A+ UBC:與已知的GRM1+ UBC(谷氨酸能)特征相似;2. CNTNAP5+ UBC:可能為未成熟的CALB2+ UBC,高表達膽堿能突觸通路相關(guān)基因。EOMES和PBX3可能是控制所有UBC譜系分化的核心調(diào)節(jié)劑,而SP5可能對CNTNAP5+譜系的發(fā)展至關(guān)重要,且具有物種保守性。
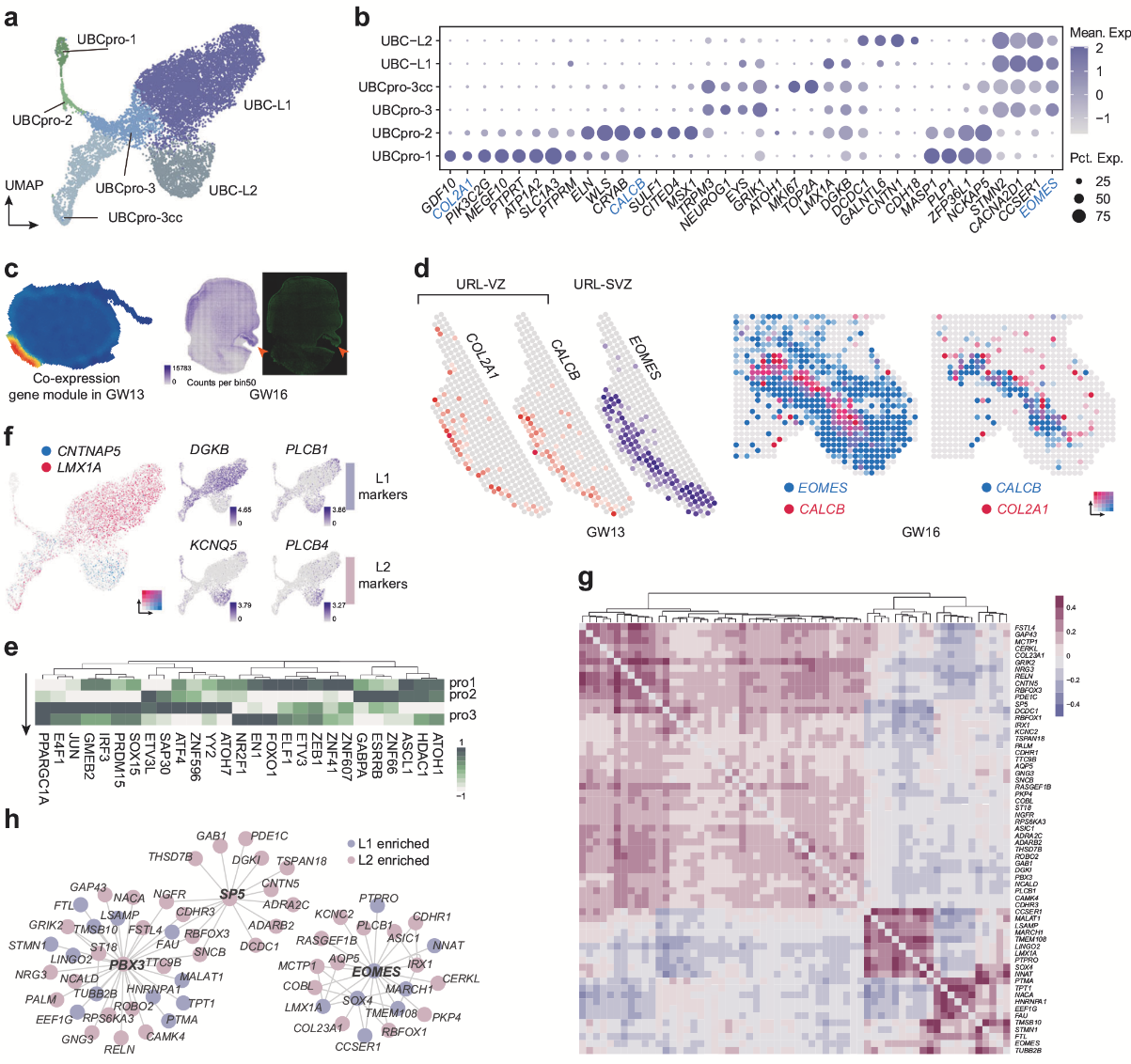
圖4 UBC祖細胞和UBC亞型的調(diào)控網(wǎng)絡(luò)
5. VZ祖細胞與中間神經(jīng)元發(fā)育
VZ可以產(chǎn)生多種類型的GABA能神經(jīng)元,如PKC。除神經(jīng)元外,VZ還生成非神經(jīng)元細胞,如OLIG2+PDGFRA+少突膠質(zhì)祖細胞,PIFO+RSPH1+室管膜祖細胞,Bergmann膠質(zhì)細胞等。PKC在小腦回路中執(zhí)行關(guān)鍵功能,本研究發(fā)現(xiàn)3種PKC分子亞型:PKC1:表達EBF2和IGF1,對應(yīng)已知的Zebrin II陰性亞型;PKC2:表達LSAMP、PCP4L1;PKC3:表達SLA、PBX3和FGF3。作者推測PKC2與PKC3均屬于Zebrin II陽性亞型。POSTN(促進神經(jīng)突生長)和IGSF21(調(diào)控抑制性突觸分化)沿A-P軸呈互補分布,暗示小腦前后區(qū)域PKCs的功能分化或發(fā)育差異。早期發(fā)育中CADM1(腹側(cè)富集)與PLCB4/CADM2(背側(cè))沿D-V軸分離(人類GW13和小鼠E15.5均存在),但后期(GW16)逐漸模糊。此外CADM1作為自閉癥相關(guān)基因,其缺失可能導(dǎo)致小腦體積縮小及PKC突觸減少,提示其在早期PKC空間組織中的潛在作用。整合scRNA-seq與scATAC-seq數(shù)據(jù)發(fā)現(xiàn),EBF2通過遠端增強子激活SEMA3A,與小鼠中Ebf2缺失導(dǎo)致Sema3a+ PKCs減少的結(jié)果一致,而NR2F2和ZNF664可能參與不同PKC亞型的分化調(diào)控。
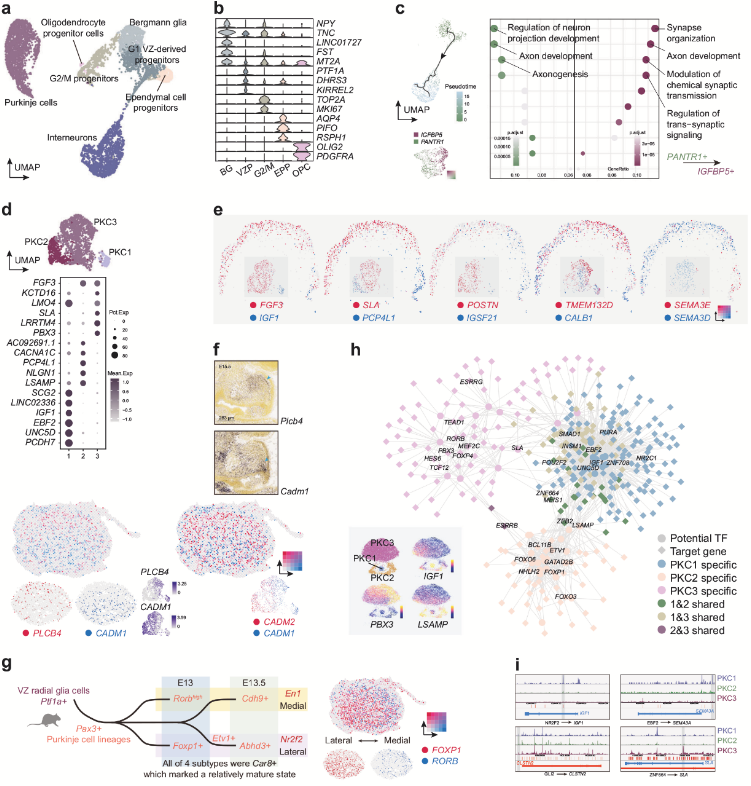
圖5 中間神經(jīng)元譜系和PKC亞型的分子特征
參考文獻:
[1] Yang F, Zhao Z, Zhang D, Xiong Y, Dong X, Wang Y, Yang M, Pan T, Liu C, Liu K, Lin Y, Liu Y, Tu Q, Dang Y, Xia M, Mi D, Zhou W and Xu Z 2024 Single-cell multi-omics analysis of lineage development and spatial organization in the human fetal cerebellum Cell Discov 10 22.