氨基酸代謝紊亂與2型糖尿病的發生密切相關。多個大規模代謝組學研究已發現,血漿中支鏈氨基酸(BCAAs)、芳香族氨基酸(AAAs)以及賴氨酸、丙氨酸、谷氨酸等水平升高與2型糖尿病風險顯著相關。然而,氨基酸如何具體調控糖代謝,尤其是其代謝產物如何通過表觀遺傳機制影響肝臟糖異生,目前尚不清楚。此外,組蛋白酰化(如乙酰化、巴豆酰化)作為連接代謝與基因表達的重要方式,近年來受到廣泛關注,但其在氨基酸代謝與糖穩態中的作用仍需深入探索。
近期,上海交通大學附屬瑞金醫院研究團隊在Advanced Science上在線發表了題為“Hepatic CBP/p300 Orchestrate Amino Acid-Driven Gluconeogenesis through Histone Crotonylation”的研究文章,結合人群遺傳學、動物模型、代謝組學、轉錄組學分析等,系統解析了CBP/p300((CREB Binding Protein,是多功能的轉錄輔激活子, 它們參與細胞周期調控、細胞分化和細胞凋亡等多種生理過程)在氨基酸驅動的糖異生過程中的關鍵作用,肝臟特異性敲除CBP/p300導致小鼠血漿氨基酸升高、氨基酸驅動的糖異生受損,關鍵機制在于氨基酸代謝基因啟動子區組蛋白巴豆酰化水平下降。在糖尿病小鼠模型中,靶向抑制CBP/p300可有效改善高血糖,為治療2型糖尿病提供了新靶點。(麥特繪譜提供Q300全定量代謝組學檢測服務)
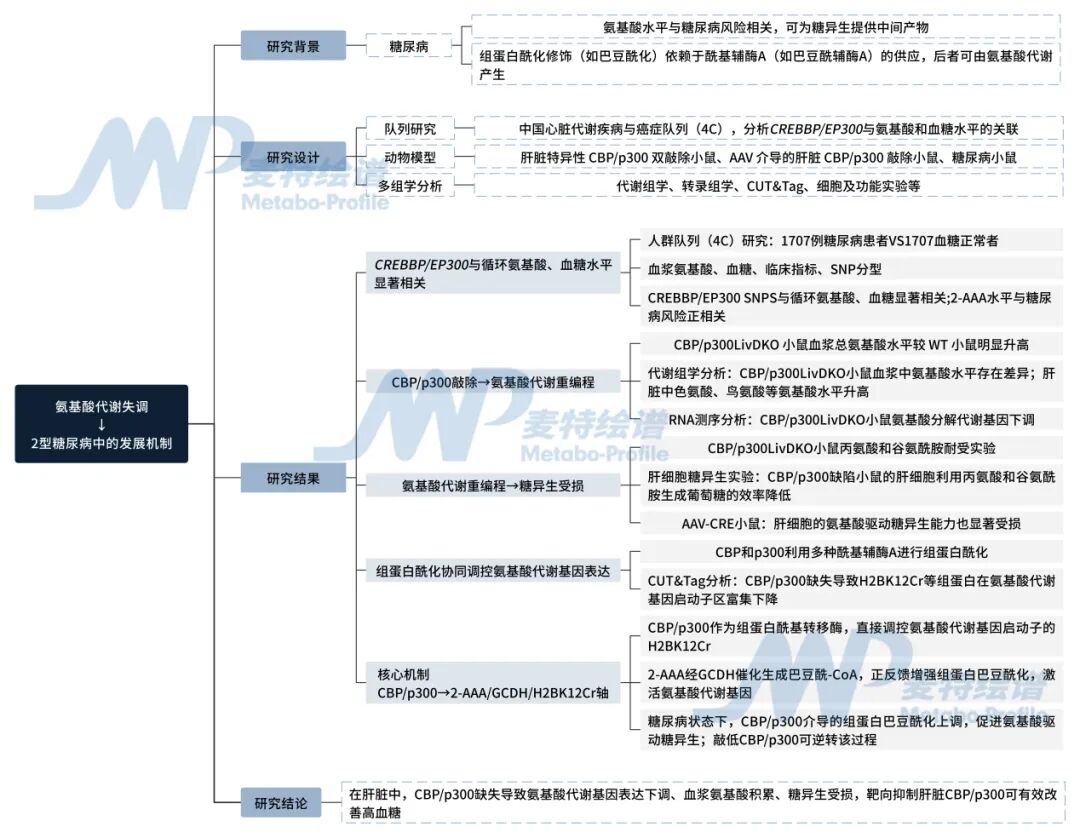
圖1.技術路線圖
研究結果
1. CREBBP/EP300基因多態性與氨基酸和血糖水平相關
CBP和p300作為組蛋白乙酰轉移酶和轉錄共激活因子,對代謝穩態至關重要,其基因單核苷酸多態性(SNPs)可能影響蛋白功能和表達,進而改變代謝通路及疾病易感性。在4C(中國心臟代謝與癌癥隊列)隊列研究中,共檢測到CREBBP(編碼CBP) 基因座內78個SNPs和EP300(編碼p300)基因座內36個SNPs,基因關聯分析顯示,23個特定 CREBBP SNPs與包括支鏈氨基酸(BCAAs)和芳香族氨基酸(AAAs)在內的多種循環氨基酸水平顯著相關,14個EP300 SNPs也與循環氨基酸譜密切相關;同時,CREBBP/EP300基因的遺傳變異還與葡萄糖穩態的關鍵臨床指標相關,這些結果提示 CBP/p300可能在連接氨基酸代謝與葡萄糖穩態中發揮作用。
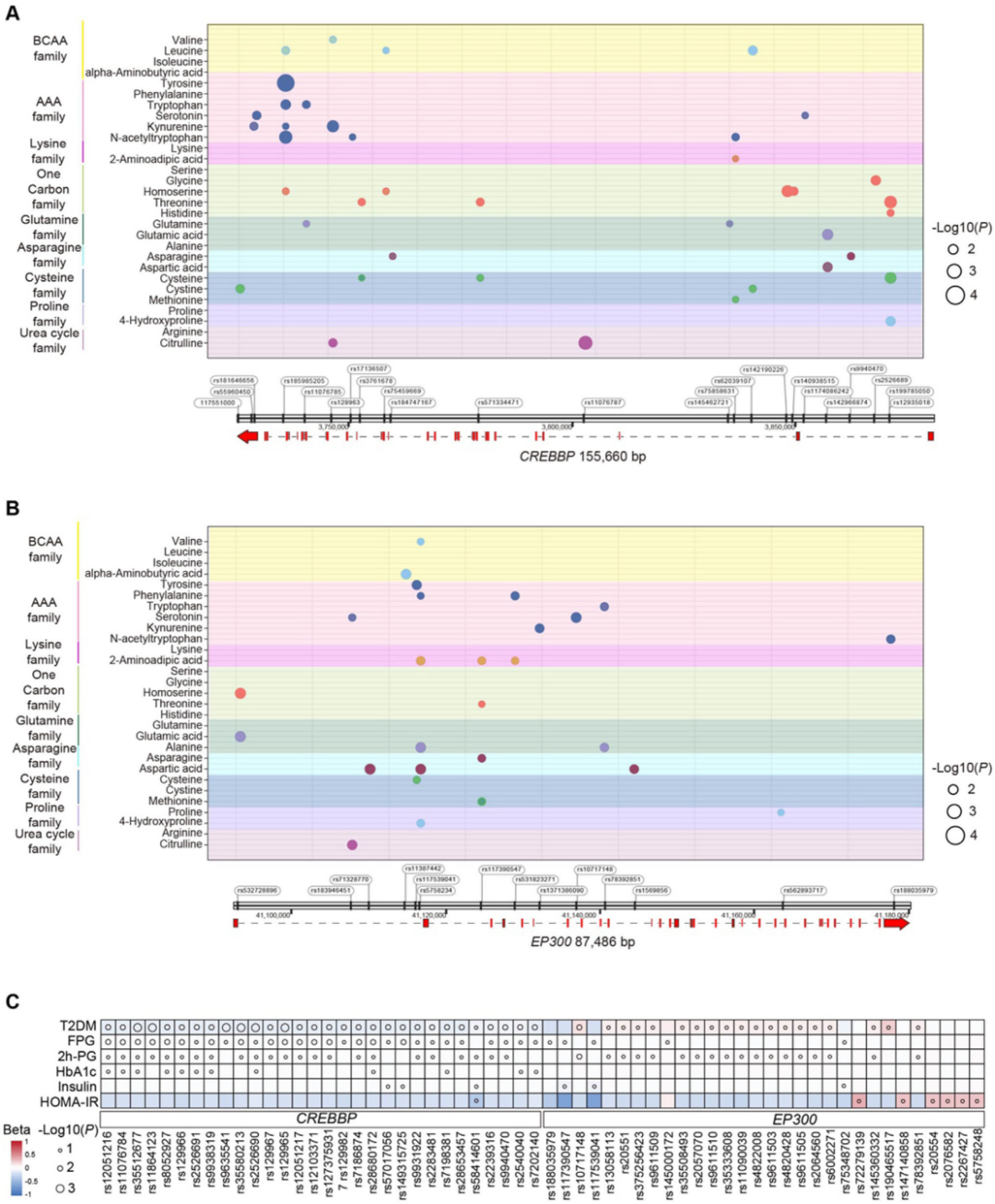
圖2.CREBBP/EP300 SNPs與循環氨基酸水平的關系
2. 肝臟Crebbp和Ep300雙敲除小鼠血漿氨基酸水平升高
鑒于肝臟在調控氨基酸和葡萄糖代謝中的核心作用,構建了多種肝臟特異性CBP/p300敲除小鼠品系,驗證肝臟中Crebbp和Ep300的敲除效果。其中,CBP/p300LivDKO (Crebbp/EP300雙敲除)小鼠空腹血糖水平顯著降低,而其他敲除品系與野生型(WT)小鼠無差異;同時,僅CBP/p300LivDKO 小鼠血漿總氨基酸水平較WT小鼠明顯升高,血漿甘油三酯(TG)、非酯化脂肪酸(NEFAs)和總膽固醇(TC)在各組間無顯著變化,這一結果證實了CBP和p300在葡萄糖和氨基酸穩態調節中的冗余功能。
代謝組學分析顯示,WT和CBP/p300LivDKO小鼠血漿代謝特征存在明顯差異,在189種已鑒定的代謝物中,差異表達代謝物主要歸類為有機酸和氨基酸,通路富集分析顯示改變的代謝物主要與氨基酸代謝通路相關,綜合單變量和多變量統計方法篩選出81個潛在生物標志物,其中包括31種氨基酸及其衍生物,20種基本氨基酸中有17種在敲除小鼠血漿中顯著升高,表明肝臟CBP/p300對維持全身氨基酸穩態至關重要。
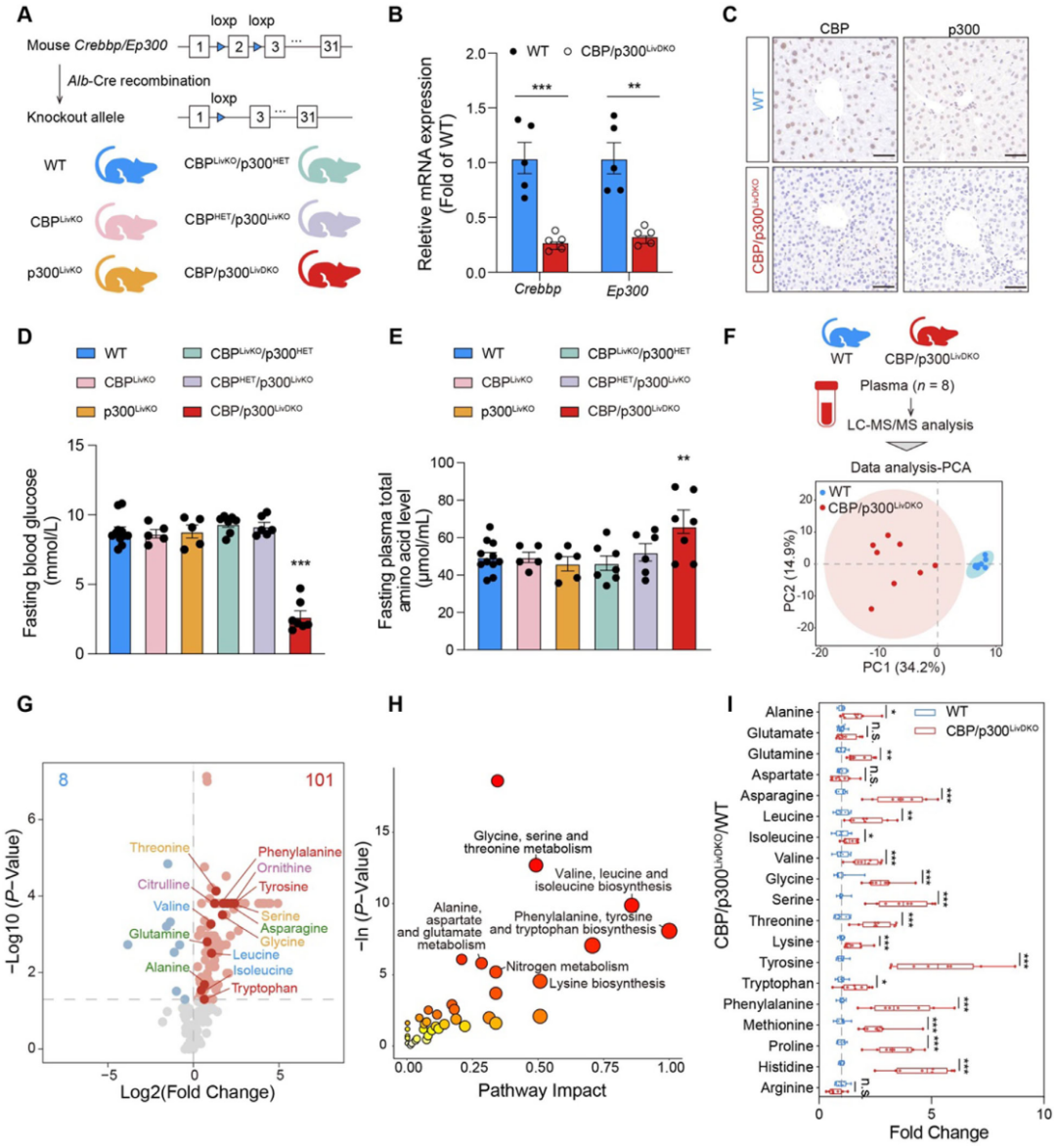
圖3.肝臟Crebbp和Ep300雙基因敲除小鼠血漿氨基酸水平升高
3. CBP/p300LivDKO小鼠肝臟氨基酸代謝重編程
為明確血漿氨基酸譜改變是否由肝臟氨基酸代謝紊亂引起,對WT和CBP/p300LivDKO小鼠肝臟進行代謝組學分析,共鑒定出213種代謝物,CBP/p300基因敲除導致肝臟中色氨酸、鳥氨酸等水平升高,支鏈酮酸(BCKAs,BCAA代謝副產物)增加但BCAA濃度與WT小鼠相當。RNA測序結果顯示,CBP/p300LivDKO小鼠肝臟中有2877個上調基因和1559個下調基因,其中多種氨基酸代謝相關基因下調,包括氨基酸分解代謝基因和氨基酸轉運體基因,但關鍵糖異生基因Pck1和G6pc表達未受影響。
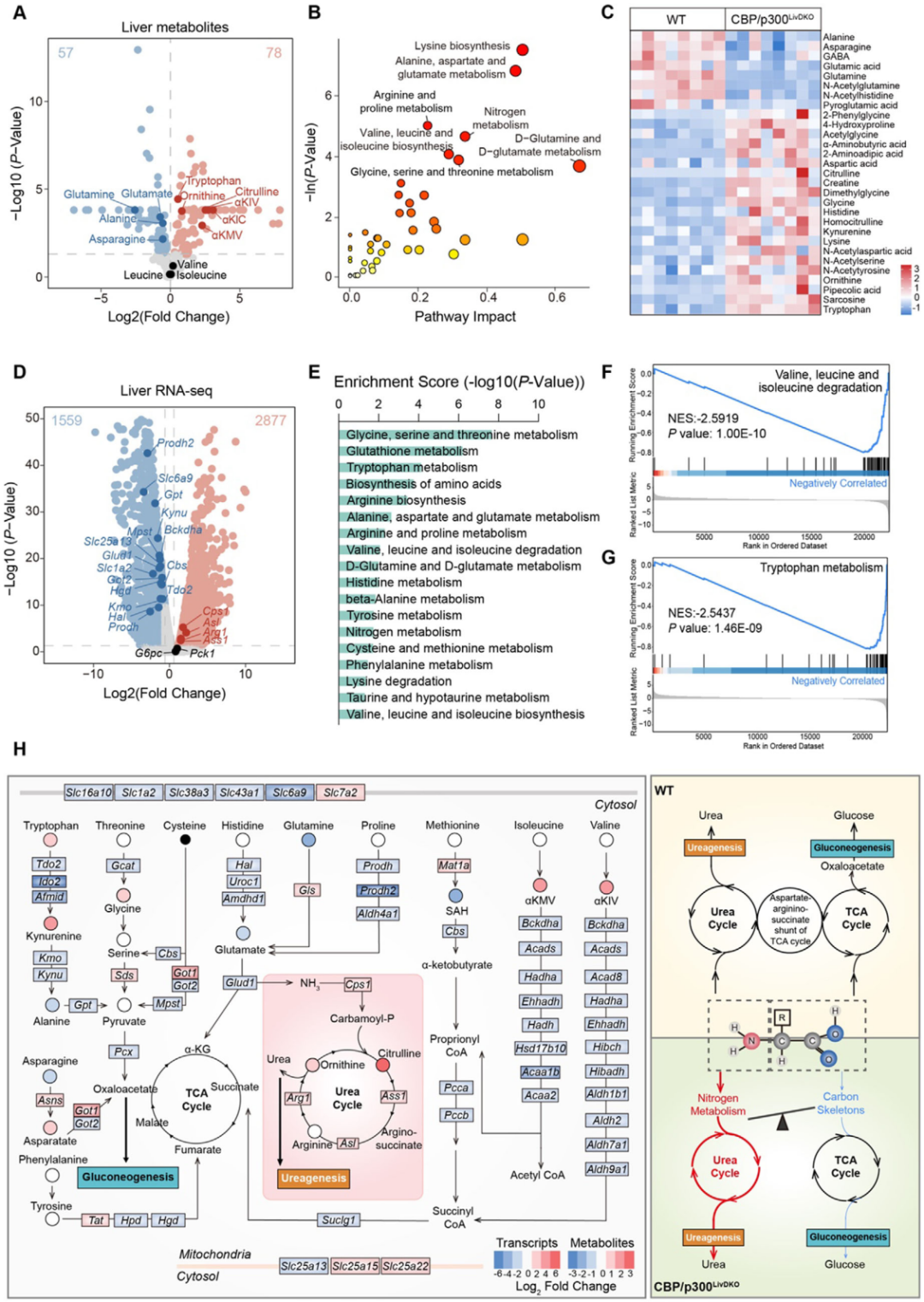
圖4. CBP/p300LivDKO小鼠肝臟氨基酸代謝重編程
4. CBP/p300LivDKO小鼠氨基酸驅動的糖異生受損
進一步探究肝臟氨基酸代謝紊亂的原因,在15種生糖氨基酸中,丙氨酸和谷氨酰胺在糖異生中起主導作用,因此對禁食小鼠進行丙氨酸耐受試驗(ATT)和谷氨酰胺耐受試驗(QTT),結果顯示,腹腔注射丙氨酸或谷氨酰胺可顯著升高 WT 小鼠血糖水平,但對CBP/p300LivDKO小鼠無明顯影響;原代肝細胞糖異生實驗進一步證實,CBP/p300缺陷小鼠的肝細胞利用丙氨酸和谷氨酰胺生成葡萄糖的效率較WT小鼠顯著降低,且伴隨多種氨基酸分解代謝基因的mRNA和蛋白水平下調。進一步驗證CBP/p300對肝臟氨基酸驅動糖異生的必要性,向Crebbpflox/flox/Ep300flox/flox小鼠注射肝臟特異性啟動子驅動的Cre重組酶腺相關病毒(AAV-CRE),3周后檢測發現,AAV-CRE小鼠肝臟中CBP/p300表達缺失,且與CBP/p300LivDKO小鼠結果一致,腹腔注射丙氨酸或谷氨酰胺后,AAV-CRE小鼠血糖水平仍維持在較低水平,其肝細胞的氨基酸驅動糖異生能力也顯著受損,同時氨基酸代謝基因的mRNA和蛋白水平降低。
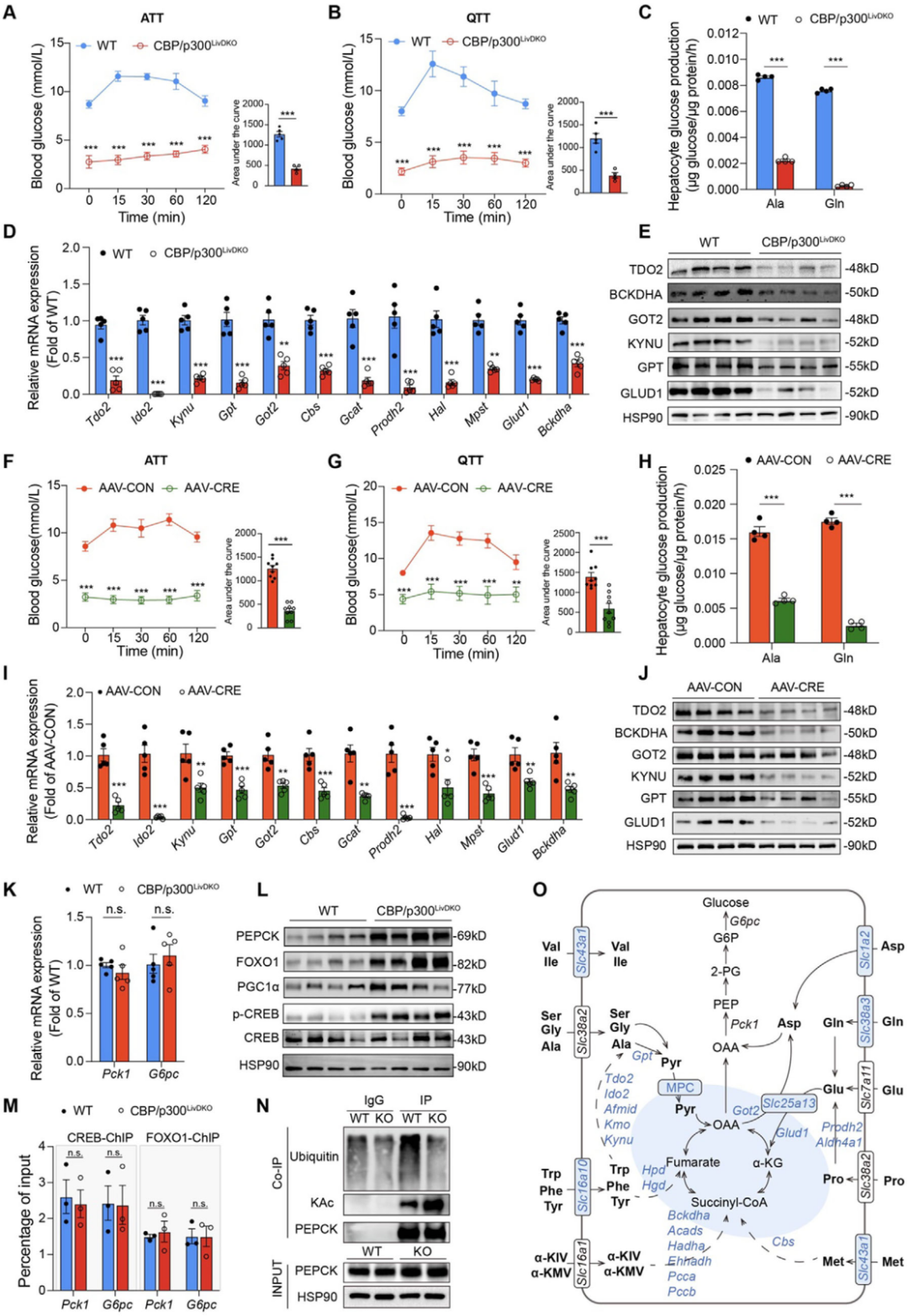
圖5.CBP/p300LivDKO小鼠氨基酸驅動的糖異生嚴重受損
5. 組蛋白酰化協同調控氨基酸代謝基因表達
CBP/p300可利用多種酰基輔酶A進行組蛋白酰化,而氨基酸分解代謝會產生乙酰輔酶A、巴豆酰輔酶A 等多種酰基輔酶A,為CBP/p300介導的組蛋白修飾提供底物,進而調控基因表達。Western blot分析顯示,CBP/p300LivDKO小鼠肝臟中,組蛋白 H2BK12Ac、H3K27Ac和H2BK12Cr水平較WT小鼠顯著降低,此外,CUT&Tag分析也顯示,CBP/p300缺失導致H2BK12Cr、H3K27Ac和H2BK12Ac在多個氨基酸代謝基因啟動子區富集程度下降,表明CBP/p300通過組蛋白修飾在連接氨基酸代謝與基因表達中發揮關鍵作用。
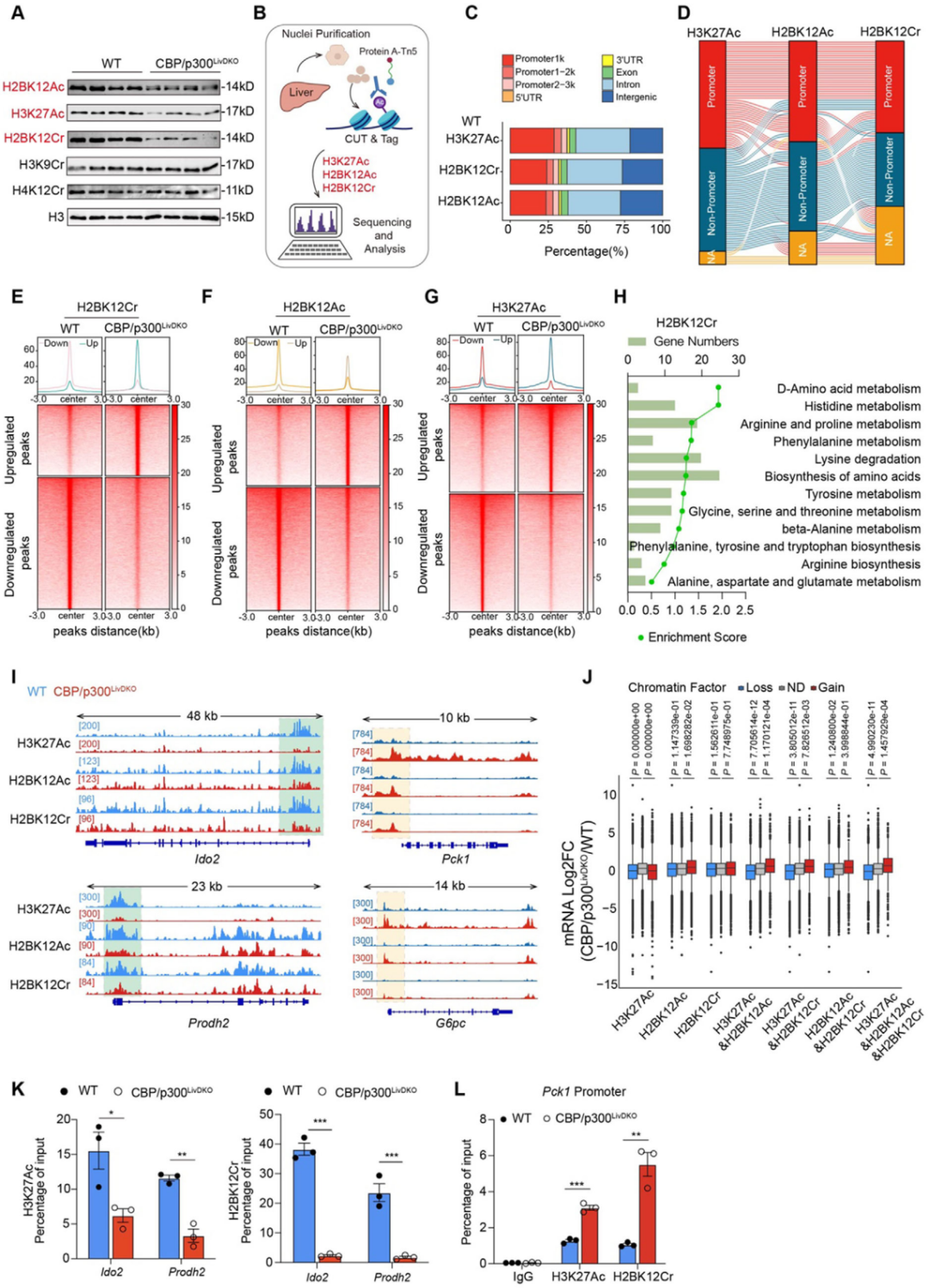
圖6. 組蛋白酰化在調節氨基酸代謝基因表達方面起協同作用
6. 2-AAA/GCDH/H2BK12Cr軸正向調控氨基酸代謝基因表達
賴氨酸降解產生的2-AAA(糖尿病生物標志物2-氨基己二酸)可在戊二酰輔酶A脫氫酶(GCDH)催化下轉化為巴豆酰輔酶A,為組蛋白巴豆酰化修飾提供底物。在原代小鼠肝細胞中加入100μM 2-AAA 處理,發現2-AAA可顯著增強由丙氨酸、谷氨酰胺和乳酸/丙酮酸驅動的糖異生,同時顯著上調 Gcdh、Prodh和Pck1的表達,且Prodh和Pck1啟動子區域的H2BK12Cr水平明顯升高。而Gcdh敲低顯著抑制由丙氨酸、谷氨酰胺和乳酸/丙酮酸介導的肝臟糖異生,同時降低Prodh和Pck1基因的表達;Gcdh沉默的肝細胞中Prodh和Pck1啟動子區域的H2BK12Ac水平顯著降低;過表達Gcdh,可部分恢復由丙氨酸和谷氨酰胺驅動的糖異生能力。
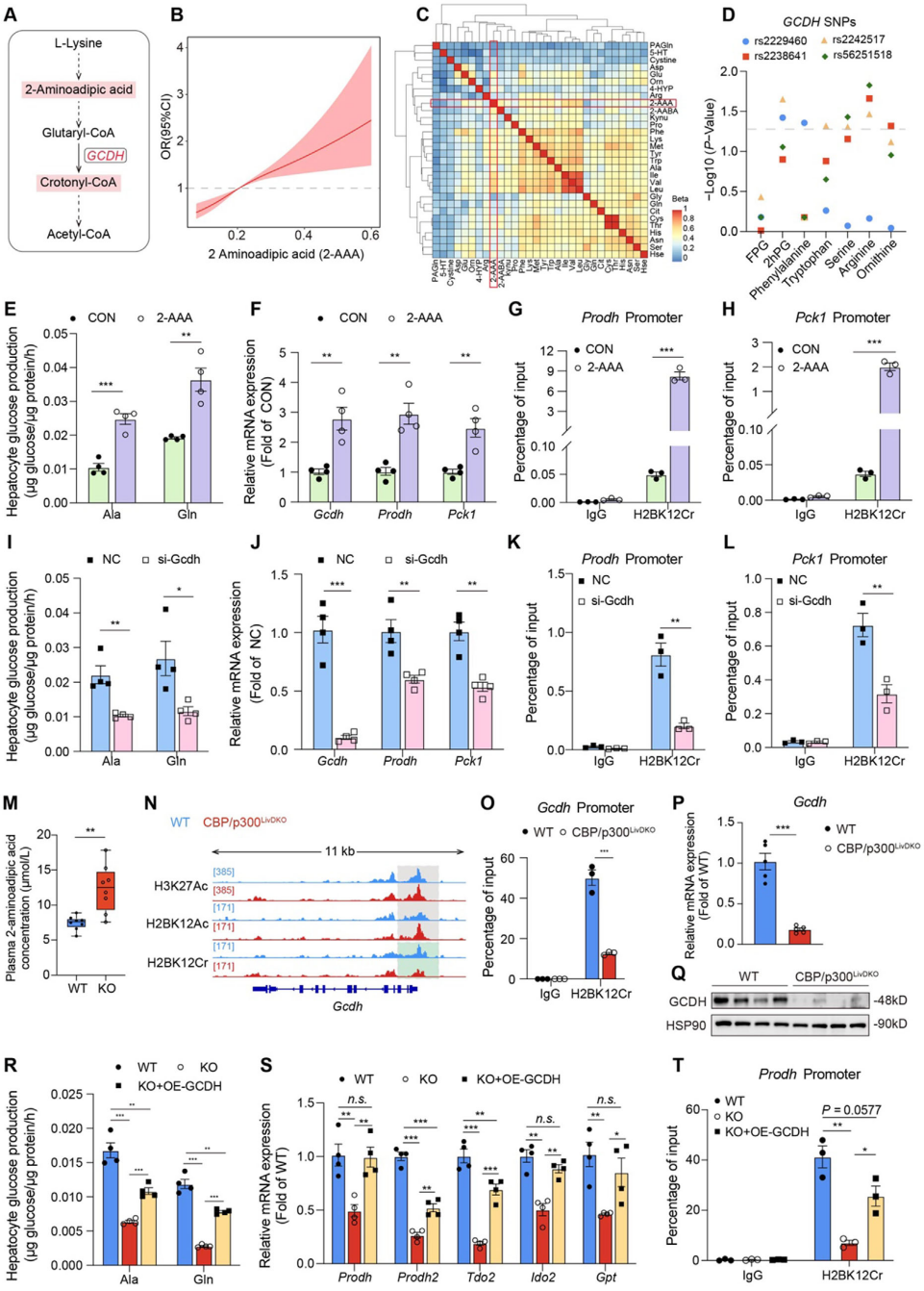
圖7.2-AAA/GCDH/H2BK12Cr軸正向調控肝臟氨基酸代謝基因的表達
7. 靶向抑制CBP/p300改善糖尿病小鼠血糖穩態
在糖尿病小鼠中,肝臟CBP/p300敲低顯著降低丙氨酸和谷氨酸誘導的血糖升高,抑制氨基酸代謝相關基因表達,并減少H2BK12Cr在關鍵基因啟動子區域的富集,進一步論證了CBP/p300是糖尿病狀態下氨基酸驅動糖異生的關鍵調控因子。
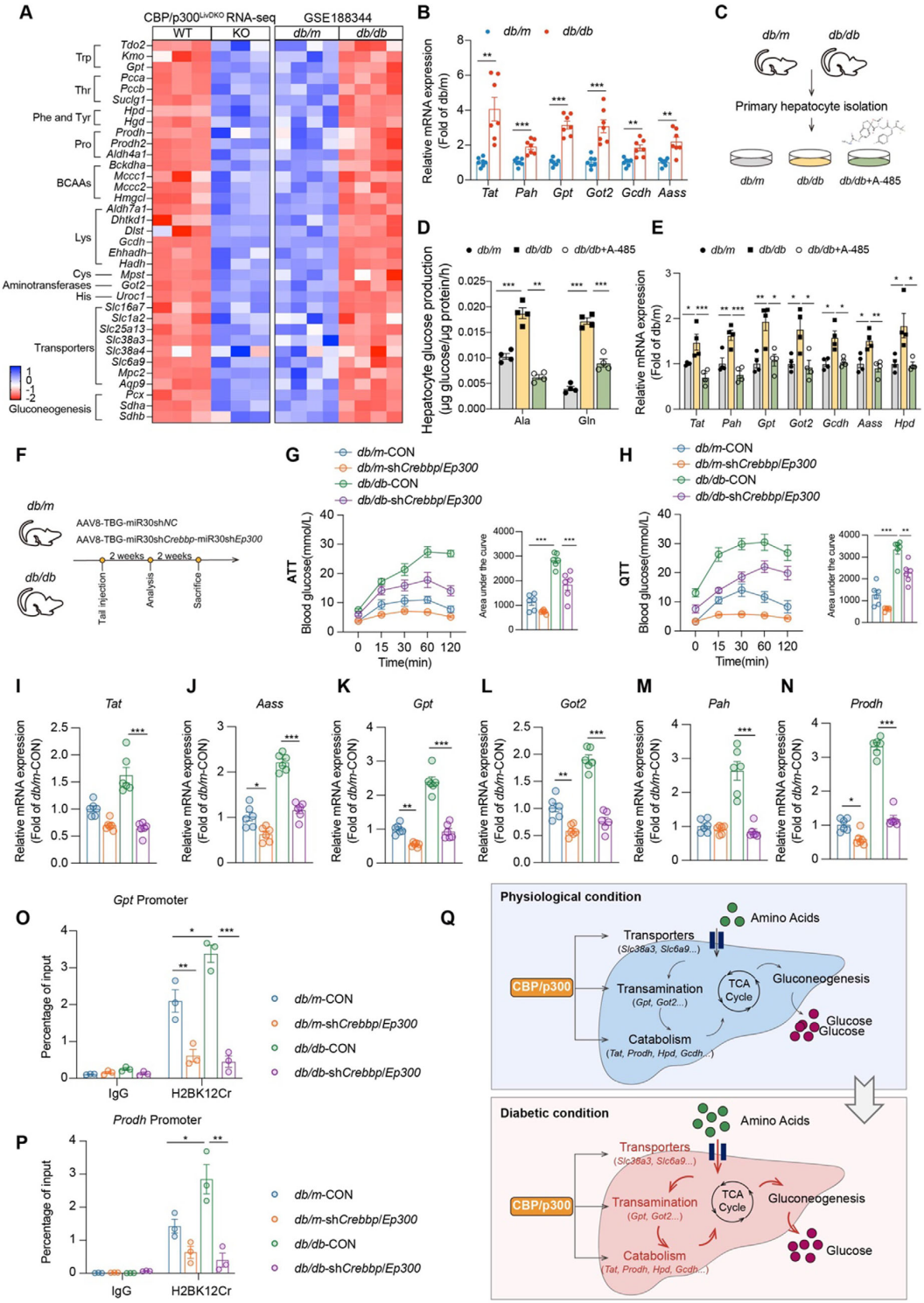
圖8.糖尿病小鼠肝臟氨基酸代謝上調
研究結論
本研究揭示了CBP/p300通過組蛋白巴豆酰化調控氨基酸代謝驅動糖異生的新機制。在肝臟中,CBP/p300缺失導致氨基酸代謝基因表達下調、血漿氨基酸積累、糖異生受損,而2-AAA/GCDH軸通過增強組蛋白巴豆酰化正向調控該過程。在糖尿病模型中,靶向抑制肝臟CBP/p300可有效改善高血糖,為2型糖尿病的干預提供了新的潛在靶點。
參考文獻
Hepatic CBP/p300 Orchestrate Amino Acid-Driven Gluconeogenesis through Histone Crotonylation. Advanced Science. 2025
繪譜幫你測
本研究前期通過人群隊列數據分析鎖定CREBBP/EP300基因多態性與氨基酸和血糖水平之間的關聯,進一步通過代謝組學技術(麥特繪譜提供Q300全定量檢測服務)對血漿和肝臟中的代謝物精準量化,證實了CBP/p300缺失對氨基酸分解代謝的影響,為后續分子機制解析“CBP/p300如何調控氨基酸代謝-糖異生通路” 提供了關鍵的代謝表型依據。
麥特繪譜開創性地搭建了醫學領域高端代謝組學技術平臺,覆蓋了非靶向-全定量-代謝流等全方位的高端醫學代謝組解決方案,同時全面布局微生物組學、轉錄組學和蛋白質組學等多組學技術服務,已成為全球多組學研究者的優選合作伙伴。麥特繪譜已為數百家三甲醫院、科研院所和企業提供多組學一站式整體解決方案,協助客戶與合作伙伴發表SCI文章600+篇,累計影響因子6000+,平均IF>10,涵蓋Cell, Science, Nature, Cancer Cell, Signal Trans-duction and Targeted Therapy, Nature Biotechnology, Cell Metabolism等權威期刊。