類器官來源于成體干細胞或多能干細胞,可分化為多種細胞類型,形成類似體內器官的結構。將類器官與微生物進行共培養,可以在體外重現宿主 - 微生物的相互作用,有助于深入理解微生物在健康和疾病中的作用機制。例如,腸道類器官可以與腸道微生物共培養,研究腸道菌群對腸道上皮細胞的影響,以及腸道上皮細胞對微生物的調控作用。
今天小編整理了一些關于類器官與微生物共培養的問題,希望能夠對各位小伙伴有所幫助。
類器官可以與多種微生物進行共培養。
細菌:結直腸癌類器官與具核梭桿菌(Fusobacterium nucleatum, Fn)共培養后,可驗證Fn的代謝產物(NaB)加強了PD-1抑制劑對腫瘤的殺傷效果[1];
病毒:Calvin J. Kuo等人[2]建立了頂端朝外的遠端肺臟類器官,其暴露的外表面上表達ACE2促進了SARS-CoV-2感染,可作為人類遠端肺部感染(包括與COVID-19相關肺炎)的體外類器官模型;
真菌:煙曲霉分生孢子或其細胞壁成分通過PRR信號通路觸發肺類器官的強烈炎癥反應,可為真菌性肺炎的機制研究提供模型[3];
寄生蟲:隱孢子蟲可成功感染并定植于人小腸與肺來源的上皮類器官中,該類器官模型能支持寄生蟲的侵入與存活,模擬生理性感染過程[4]。
目前,關于微生物感染類器官的方法主要有以下幾種:
可以。目前在疫苗研發應用最廣的是扁桃體類器官,利用類器官樣本與流感等抗原共同孵育,結合基線血液和組織多維度測試,分析不同時間點免疫細胞組成和激活、抗體特異性以及中和能力等[17]
1. 為了確保微生物能在適宜的環境下與類器官進行共培養,需要避免添加抑制其生長的抗生素;
2.對于厭氧微生物的共培養,需要調整氧氣濃度[5];
3.根據微生物的需求,可額外補充碳源/氮源或其他代謝底物。
1、用熒光染料對微生物進行染色,有些可以追蹤它們在共培養中的行為長達1周[5]。
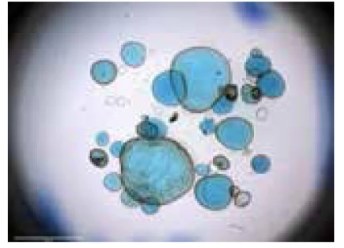
圖1. 顯微注射法實現微生物與類器官共培養[5]
2、也可以采用電鏡的方式直接觀察[18]。
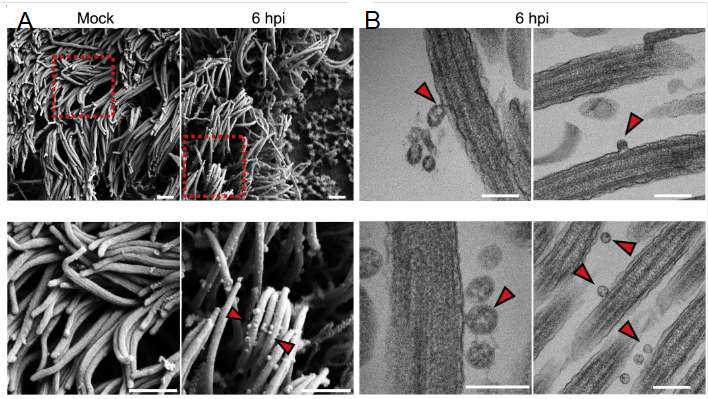
圖2. 利用SEM(A)和TEM(B)觀察被病毒感染前(Mock)和感染后6小時(6 hpi)的類器官纖毛部分,箭頭部分為病毒[18]
3、還可以將類器官提取RNA,檢測微生物的16S rRNA的表達水平,在宿主細胞的管家基因歸一化之后即可表示感染情況[6] 。
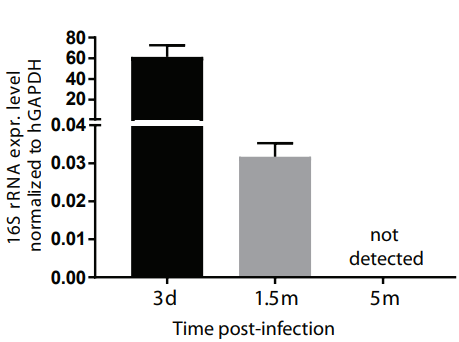
圖3. 檢測16S rRNA表達水平評估細菌感染類器官的強度[6]
4、可以利用現有的滴度檢測方法,如Rotavirus Fluorescent Focus Assay (FFA)可以檢測輪狀病毒的產量[19]。
大體流程是一樣的,只是類器官的培養體系可能需要額外的操作步驟,如:對于非分泌型微生物,如果宿主類器官是包埋在基質膠內培養的,需要先將基質膠消化后,再進行收集或提取。
有這樣的微生物,如:諾如病毒(Norovirus, NV)。諾如病毒于1972年由Kapikian等在美國諾瓦克市腹瀉暴發事件中首次分離,NV只能使用類器官培養主要是有以下幾個原因:
1、該病毒可在腸上皮細胞中檢測到,但在傳統細胞系中難以穩定復制;
2、該病毒具有高度物種專一性,無法通過常規實驗動物模擬人類感染;
3、而經過實驗驗證,該病毒可以感染腸道類器官并且實現有限復制[20]。
基于類器官獨特的3D結構,有些實驗是無法用二維細胞完成的:
1、特定微生物的體外擴增只能在類器官中完成,如上文提到的諾如病毒[20];
2、類器官的三維結構可以用于模擬體內的微生物定植環境,進行微生物定植相關的研究[21];
3、細胞系的基礎性質與它們來源的細胞有很大差異,如:肝細胞系與其相關的原代細胞進行比較時,觀察到轉錄組學和蛋白質組學表達的偏差[22],可能影響感染數據的準確性;
4、由于類器官的藥敏測試與臨床結果更接近,因此在抑菌/抑制病毒等藥物的研發、以及引入免疫細胞后的免疫效果評估會更準確。
感染前的微生物最好做一些數量及活性的評估,以下步驟可按需選擇:
1、確保微生物結構的完整性;
2、確保微生物的活力:細菌在感染前可以先做生長曲線,最好在對數期時使用;真菌可選擇鈣熒光白(CFW)染色確認活性;寄生蟲可以在顯微鏡下確定數量與發育階段,并在鏡下觀察蠕動或游動;
3、預實驗確定MOI,之后可按需選擇濃縮步驟。
Toshiro Sato等人[23]開發了一種名為 iHACS(腸道半厭氧共培養系統)的新技術,它可以模擬人類結腸上皮細胞和腸道厭氧菌的共生環境。該系統包含一個低氧的頂端室和一個常氧的基底室,允許厭氧菌在低氧環境下生長,同時維持上皮細胞的活力。
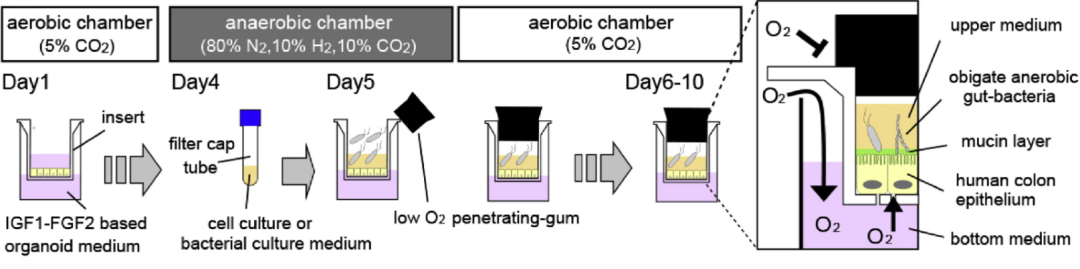
圖4. 使用Transwell的腸類器官和厭氧微生物群的共培養體系示意圖[23]
? 相關產品
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
參考文獻
[1]Wang, X. et al. (2024). Fusobacterium nucleatum facilitates anti-PD-1 therapy in microsatellite stable colorectal cancer. Cancer Cell, 42(10), 1729-1746.
[2]Salahudeen, A. et al. (2020). Progenitor identification and SARS-CoV-2 infection in human distal lung organoids. Nature, 588(7839), 670-675.
[3]Bosáková, V. et al. (2023). Activation of TLRs by opportunistic Fungi in lung organoids. In Toll-Like Receptors: Methods and Protocols (pp. 271-284). New York, NY: Springer US.
[4]Heo, I. et al. (2018). Modelling Cryptosporidium infection in human small intestinal and lung organoids. Nature microbiology, 3(7), 814-823.
[5]Puschhof, J. et al. (2021). Intestinal organoid cocultures with microbes. Nature protocols, 16(10), 4633-4649.
[6]Kessler, M. et al. (2019). Chronic Chlamydia infection in human organoids increases stemness and promotes age-dependent CpG methylation. Nature communications, 10(1), 1194.
[7]Stroulios, G. et al (2021). Culture methods to study apical-specific interactions using intestinal organoid models. Journal of Visualized Experiments (JoVE), (169), e62330.
[8]Stroulios, G. et al. (2022). Apical-out airway organoids as a platform for studying viral infections and screening for antiviral drugs. Scientific reports, 12(1), 7673.
[9]Cho, Y.. et al. (2024). Establishment of an Apical-Out Organoid Model for Directly Assessing the Function of Postbiotics. Journal of Microbiology and Biotechnology, 34(11), 2184.
[10]Ahmad, V. et al. (2024). Development of polarity-reversed endometrial epithelial organoids. Reproduction, 167(3).
[11]Jenal, U. (2024). Pseudomonas aeruginosa breaches respiratory epithelia through goblet cell invasion in a microtissue model. Nature Microbiology, 9(7), 1725-1737.
[12]Aguilar, C. et al. (2021). Organoids as host models for infection biology–a review of methods. Experimental & Molecular Medicine, 53(10), 1471-1482.
[13]Sachs, N. et al. (2019). Long‐term expanding human airway organoids for disease modeling. The EMBO journal, 38(4), e100300.
[14]Horsley, H. et al. (2018). A urine-dependent human urothelial organoid offers a potential alternative to rodent models of infection. Scientific reports, 8(1), 1238.
[15]Boccellato, F. et al. (2019). Polarised epithelial monolayers of the gastric mucosa reveal insights into mucosal homeostasis and defence against infection. Gut, 68(3), 400-413.
[16]Alonso-Roman, R. et al. (2024). Organ-on-chip models for infectious disease research. Nature microbiology, 9(4), 891-904.
[17]Wagoner, Z. W. et al. (2025). Systems immunology analysis of human immune organoids identifies host-specific correlates of protection to different influenza vaccines. Cell Stem Cell, 32(4), 529-546.
[18]Wu, C. T. et al. (2023). SARS-CoV-2 replication in airway epithelia requires motile cilia and microvillar reprogramming. Cell, 186(1), 112-130.
[19]Finkbeiner, S. R. et al. (2012). Stem cell-derived human intestinal organoids as an infection model for rotaviruses. MBio, 3(4), 10-1128.
[20]Hosmillo, M. et al. (2020). Norovirus replication in human intestinal epithelial cells is restricted by the interferon-induced JAK/STAT signaling pathway and RNA polymerase II-mediated transcriptional responses. MBio, 11(2), 10-1128.
[21]McCracken, K. W. et al. (2014). Modelling human development and disease in pluripotent stem-cell-derived gastric organoids. Nature, 516(7531), 400-404.
[22]Olsavsky, K. M. et al. (2007). Gene expression profiling and differentiation assessment in primary human hepatocyte cultures, established hepatoma cell lines, and human liver tissues. Toxicology and applied pharmacology, 222(1), 42-56.
[23]Sasaki, N. et al. (2020). Development of a scalable coculture system for gut anaerobes and human colon epithelium. Gastroenterology, 159(1), 388-390.